Quello che tu chiami schiavo pensa che è nato come te, gode dello stesso cielo, respira la stessa aria, vive e muore, come viviamo e moriamo noi. Puoi vederlo libero cittadino ed egli può vederti schiavo.
Seneca
INTRODUZIONE
Il Paleolitico (dal greco palaios, antico, e lithos, pietra, ossia "età della pietra antica" o "età antica della pietra") è il periodo della preistoria in cui si sviluppò la tecnologia umana con l'avvento dei primi strumenti in pietra da parte di diverse specie di ominidi. Iniziò circa 2,5 milioni di anni fa e terminò 10.000 anni fa con l'introduzione dell'agricoltura e il passaggio al Mesolitico, o, nelle zone di precoce neolitizzazione, all'Epipaleolitico.
Il termine fu introdotto dallo studioso John Lubbock nel 1865 in opposizione al termine "Neolitico". Tra le epoche geologiche corrisponde al Pleistocene. Nella scala dei tempi geologici, il Pleistocene è la prima delle due epoche in cui è suddiviso il Quaternario. È, convenzionalmente, compreso tra 2,58 milioni di anni fa e 11.700 anni fa, preceduto dal Pliocene, l'ultima epoca del precedente periodo del Neogene, e seguito dall'Olocene, l'epoca in cui viviamo.
Giova ricordare che il mesolitico o epipaleolitico è il periodo intermedio dell'Età della pietra che va dal 10.000 all'8.000 a.C., definito per questo l'età della pietra di mezzo. Durante il Mesolitico si elaborano tecniche sofisticate di lavorazione della pietra, come quella della "microlitica", nella quale piccole schegge di selce fissate a manici in legno o in osso sono utilizzate per costruire utensili per la caccia e la raccolta dei vegetali. Si ha inoltre uno sviluppo delle armi da lancio e in particolare si generalizza l'impiego dell'arco e della freccia, soprattutto in Europa. Ciò è dovuto a rilevanti cambiamenti riguardanti il clima; infatti il mesolitico si estende nel periodo finale dell'ultima glaciazione, dopo l'ultimo massimo glaciale e fino all'instaurazione di un clima simile a quello attuale. Il cambiamento climatico fu causa o concausa della scomparsa di grandi animali come il mammut e i gliptodonti, favorì l'espansione di vaste foreste al posto delle tundre e conobbe una crescita demografica dell'umanità primitiva senza precedenti. Le abitazioni sono costituite da capanne che formavani i villaggi.
La grotta di Chauvet
La grotta Chauvet è uno dei più noti e importanti siti europei, ricco di testimonianze, simboliche ed estetiche, del Paleolitico superiore. È il più antico esempio di arte preistorica del mondo.
La grotta Chauvet, che prende il nome dal suo scopritore, si trova presso la colline delle Cirque d'estre Vallon-Pont-d'Arc nell'Ardèche. La grotta presenta pitture e incisioni di diversi animali quali bisonti, mammut rossi, gufi, rinoceronti, leoni, orsi, uri, cervi, cavalli, iene, renne, lupi (pochi) ed enormi felini scuri (forse leopardi). Soli o ritratti in branco, nei colori resi disponibili dagli elementi naturali, gli animali ritratti assommano ad oltre 500 opere databili dai 32.000 ai 36.000 anni fa, nel Paleolitico superiore. Possiamo ipotizzare che questo luogo fosse un importante centro di culto dell'epoca. Le figure hanno un dinamismo potente e la mancanza di definizione contribuisce a dare all'insieme un carattere magico. Gli animali paiono uscire dalla roccia stessa o rientrarvi a seconda della prospettiva e dei giochi di luce. Questo tipo di organizzazione estetica è comune a tutta l'arte visiva del Paleolitico. Nella grotta troviamo anche ossa di vari animali, teschi di Ursus spelaeus e una roccia che per risalto e forma può sembrare quasi un altare primitivo, su cui è appoggiato un teschio di orso, quasi fosse stato offerto o utilizzato per qualche rito.
Se si accetta questa teoria, la sala più interna della grotta (la Sala del Fondo), avrebbe potuto rappresentare il fulcro del viaggio iniziatico. Qui infatti scende un pendente che arriva a 1,20 m dal suolo, e da questo, uno strano essere ibrido e antropomorfo guarda la parete dinnanzi. Ha corna frontali che gli danno il vago aspetto di un bisonte e occhi tondi come se fossero di un insetto. Alcuni lo hanno chiamato lo Stregone della grotta.
Il pendente ha una chiarissima forma fallica e si pone davanti a una cavità dall'evidente forma vaginale. Le forme del pendente, incompiute e difficili da analizzare, paiono tracciare le linee opulente di un corpo femminile, molto simili alle statuette a tutto tondo reperite in numerosi luoghi in Europa.
Questa suggestiva fusione di forme maschili e femminili, animalesche e umane, aiutata dalle caratteristiche proprie della "sala" stessa deve averle dato una valenza particolare. Una terza figura unisce la donna all'essere ibrido ed è una chiara figura leonina, che ancora rimanda al cerchio morte-vita-rigenerazione che emerge chiaramente in tutte le testimonianze di questo nostro passato ancestrale. La figura del leone diverrà una figura mitica e allegorica sin nei bestiari medioevali, restando associata ai suoi significati originari. Il leone impersona la figura di un Dio arbitro di morte e portatore di vita, ma soprattutto di una rinascita totale che è fuori dalle possibilità dell'uomo. Questa ipotesi suggestiva si basa sull'analisi comparata di forme religiose arcaiche sopravvissute sino ad oggi o documentate (a partire da Frazer, ne Il ramo d'oro) più che su fatti archeologici, ritrovamenti o prove. La realtà è che non sappiamo nulla di una cultura che definita appunto "pre-istorica", non ha lasciato molto di sé. Ipotesi per ipotesi, altrettanto suggestiva potrebbe essere la teoria secondo cui un individuo asociale, magari emarginato o pazzo, sia penetrato nella grotta e suggestionato dal luogo abbia incominciato a dipingere. Non ne sappiamo abbastanza, e dobbiamo tenerci lontani dal confondere le ipotesi dai fatti. Quello che emerge dai "fatti" è solo che vi sono molte pitture eseguite da uno stesso individuo (riconoscibile perché c'è traccia di un suo lieve difetto fisico: il quinto dito della mano destra presenta la falangina lievemente piegata verso l'interno), che molte pitture hanno al di sotto tracce di immagini precedenti anche di 5.000 anni, che non vi sono resti umani, che la grotta una volta era meno profonda e disagevole (si è poi sigillata a causa di una frana), che vi venivano adoperate torce in legno, che era frequentata da orsi non si sa se contemporaneamente alla frequentazione umana, che le pitture paiono così fresche perché ricoperte di calcite. La grotta è ricca di evoluzioni nella tecnologia della lavorazione della pietra, dei materiali scolpibili (steatite, avorio) e nella produzione di nuove armi e tecniche di raccolta. È caratterizzata da un'arte volta alla scultura e all'incisione di figure femminili, solitamente in pietra o avorio, di gioielli e soprattutto di ornamenti, simbolo di appartenenza a un credo e a un gruppo sociale. Questi sono usualmente fatti di conchiglie, denti di animali e steatite. I reperti di questo periodo sono altamente figurativi e spesso complessi, grazie a una struttura sociale che, supponiamo, era in grado di comunicare, tramandare e trasporre visivamente concetti simbolici articolati che caratterizzeranno l'opera umana in tutti i periodi a venire. I materiali usati in queste lontane forme d'arte sono moltissimi e scelti con cura, come con cura erano lavorati. Ulteriori reperti ci dicono chiaramente che la musica faceva già parte dell'espressività di quei popoli.

Gruppo di cavalli, disegnato nella grotta Chauvet
La grotta di Altamira
La grotta di Altamira è una caverna spagnola famosa per le pitture parietali del Paleolitico superiore raffiguranti mammiferi selvatici e mani umane. Si trova nei pressi di Santillana del Mar in Cantabria. È stata inclusa tra i Patrimoni dell'umanità dell'UNESCO nel 1985. Nel 2008 il nome del patrimonio è stato modificato da "Grotta di Altamira" in "Grotta di Altamira e arte rupestre paleolitica della Spagna settentrionale" in seguito all'aggiunta di 17 altre grotte.
La grotta originaria è lunga 270 metri e consiste di una serie di passaggi intrecciati e di camere. La caverna si è formata grazie al crollo di precedenti fenomeni carsici nella roccia calcarea del monte Vispieres.
Gli scavi archeologici nel fondo della cava hanno portato alla luce ricchi depositi di arte del Solutreano superiore (circa 18.500 anni fa) e del Magdaleniano inferiore (tra i 16.500 e i 14.000 anni fa). Nel lungo intervallo di tempo fra questi due periodi di occupazione umana la grotta è stata usata da animali selvatici. Il sito si trova in un punto strategico per poter sfruttare la disponibilità di cibo costituito dalla ricca fauna che abitava le vallate delle montagne circostanti. Circa 13.000 anni fa una frana bloccò l'entrata della caverna, preservandone così il contenuto fino alla scoperta avvenuta nel 1879 in seguito al crollo di un albero.
L'occupazione umana della grotta è stata limitata all'entrata, nonostante siano state trovate pitture per tutta la lunghezza del cunicolo. Gli artisti usarono carboncino e ocra o ematite per dipingere, spesso diluendo i colori per produrre tonalità diverse e creare così effetti di chiaroscuro; sfruttarono anche i contorni naturali dei muri per dare un'impressione di tridimensionalità ai soggetti. Il Soffitto Multicolore è l'opera più appariscente e mostra un branco di bisonti in differenti posizioni, due cavalli, un grande cervo e probabilmente un cinghiale.
La datazione di queste opere si può far risalire al Magdaleniano per quanto riguarda animali e forme astratte. Le immagini attribuite al Solutreano raffigurano invece cavalli, capre e impronte di mani. Numerose altre caverne nel nord della Spagna contengono esempi di arte paleolitica, ma nessuna di loro è qualitativamente o quantitativamente paragonabile ad Altamira.
Nel 1879 l'archeologo dilettante Marcelino Sanz de Sautuola scoprì le pitture sulla volta della grotta. La grotta venne poi scavata dallo stesso Sautuola e dall'archeologo Juan Vilanova y Piera dell'Università di Madrid che riportarono i risultati del loro lavoro in un notissimo studio pubblicato nel 1880 in cui facevano risalire all'età paleolitica le opere rinvenute. Gli specialisti francesi, guidati da Gabriel de Mortillet ed Émile Cartailhac, rigettarono senza appello le ipotesi di Sautuola e Piera e le loro scoperte furono ridicolizzate al Congresso preistorico di Lisbona del 1880. A causa della loro elevata qualità artistica e dell'eccezionale stato di conservazione, Sautuola venne anche accusato di truffa; un contadino locale sostenne addirittura che le pitture erano state create da un artista contemporaneo su ordine di Sautuola.
Fu solo nel 1902, dopo che altre scoperte avevano contribuito ad avallare l'ipotesi dell'estrema antichità dei dipinti di Altamira, che la società scientifica rivide il proprio giudizio sulle scoperte dei due spagnoli. In quell'anno Émile Cartailhac ammise enfaticamente il proprio errore nel celebre articolo intitolato Mea culpa d'un sceptique, pubblicato sul giornale L'Anthropologie, mentre un altro archeologo francese, Joseph Déchelette, definì Altamira "la Cappella Sistina della preistoria". Sautuola, morto 14 anni prima, non poté assistere al trionfo delle sue teorie.
Iniziarono immediatamente altri scavi, effettuati da Hermilio Alcalde del Río (1902-1904), cui seguirono quelli del tedesco Hugo Obermaier (1924-1925) e, più avanti, di Joaquín González Echegaray (1981). Nel 2008, infine, grazie al sistema di datazione uranio-torio, alcuni dipinti sono stati fatti risalire a 35.000-25.000 anni fa. Inoltre, studi recenti confermano l'ipotesi che in vari casi ci si trovi di fronte a opere "collettive" completate nell'arco di migliaia di anni.

Bisonti raffigurati nel soffitto della grotta di Altamira
Ambiente in Europa durante il paleolitico
Durante il paleolitico sono avvenute una serie di glaciazioni, note come glaciazione di Günz (Günz è il nome dato alla prima glaciazione avvenuta in Europa, che ebbe luogo nel Pleistocene e in cui si verificò un abbassamento generale della temperatura e un'espansione dei ghiacciai fino all'attuale zona temperata.
Questa prima glaciazione si verificò da 1.200.000 anni fa fino a circa 900.000 anni fa, e fu seguita da un periodo interglaciale che durò circa 150.000 anni), di Mindel (E' la seconda glaciazione in Europa avvenuta nel Pleistocene in cui si verificò un abbassamento generale della temperatura e un'ulteriore espansione dei ghiacciai nell'attuale zona temperata. Questa seconda glaciazione iniziò 455.000 anni fa e durò fino a 300.000 anni fa, e fu seguita poi da un secondo periodo interglaciale che durò circa 100.000 anni. In questo periodo l'uomo iniziò ad utilizzare il fuoco), di Riss (E' la terza glaciazione in Europa avvenuta nel Pleistocene in cui si verificò un'ulteriore espansione dei ghiacciai nell'attuale zona temperata. Questa terza glaciazione iniziò 200.000 anni fa e durò fino a 130.000 anni fa, seguita poi da un periodo interglaciale che durò circa 20.000 anni) e di Würm (In realtà rappresenta l'effetto prodotto dall'ultima glaciazione su una zona specifica come le Alpi o la Sierra Nevada, ma per convenzione essa viene estesa anche a livello globale come l'equivalente di ultimo periodo glaciale, il più recente periodo glaciale incominciato circa 110.000 anni fa e terminato all'incirca 11.700 anni fa). Durante le epoche glaciali i ghiacci avevano coperto gran parte dell'Europa settentrionale e centrale, spingendosi fin quasi sulle coste del Mar Mediterraneo e provocando l'abbassamento del livello del mare di oltre 100 metri. Avvenivano quindi contatti tra gli abitanti della penisola iberica e di quella italica. Con la fine dell'ultima glaciazione, tra 15.000 e 10.000 anni fa, e il conseguente aumento delle temperature, i ghiacciai ripresero a sciogliersi, e il livello dei mari si rialzò nuovamente.
Vita nel Paleolitico
I gruppi umani, prevalentemente nomadi o a sedentarizzazione periodica, erano caratterizzati da un'economia di caccia e raccolta, che si andò evolvendo con lo sviluppo di forme di caccia specializzata e con la pesca. Alcune teorie sostengono che soprattutto le donne con i bambini andassero a raccogliere erbe, radici e frutti selvatici. Invece gli uomini organizzavano battute di caccia in gruppo per animali di grossa taglia o si dedicavano alla pesca. Le abitazioni erano inizialmente semplici ripari naturali, a cui si aggiunsero capanne costruite con pelli e ossa di animali. In questo periodo si iniziò a controllare il fuoco e poi ad accenderlo. Il fuoco venne utilizzato come protezione dagli animali, per illuminare, per cucinare e anche per riscaldarsi.
L'arte
La specie homo sapiens (vedi sotto) sembra che evolva in maniera indipendente dal neanderthal, a partire circa da 200.000 anni fa (alcune scuole parlano di 300.000 anni fa). I resti più antichi di umani indubitabilmente moderni si ritrovano al sito Kibish nei pressi del fiume Omo, in Etiopia e nel sito Qafzeh-Skhul nell'attuale Israele.
In terra africana la specie evolve culturalmente. Sono numerosi i ritrovamenti fossili e di manufatti e, a tutt'oggi, il più antico ritrovamento di un oggetto dalle indubbie caratteristiche artistiche risale a 80.000 anni fa, in prossimità di Cape Agulhas, nella Caverna di Blombos, in Sud Africa.
L'evoluzione, fisica, tecnologica e culturale, condurrà alla fine del paleolitico e all'inizio del neolitico.
Negli ultimi anni si è rafforzata la teoria, pur con altalenanze scientifiche tuttora in corso, che vede neanderthal e sapiens (tra cui la popolazione Cro-Magnon) come due specie diverse evolutesi in modo quasi parallelo. L'uomo di Cro-Magnon, ovvero ascrivibile all'uomo moderno, sostituisce in Europa l'uomo di Neanderthal (che pare si estingua circa 28.000 anni fa) in un arco di tempo relativamente breve ma con una certa sovrapposizione di alcune migliaia di anni, anche se non è ancora possibile stabilire che tipo di relazioni (collaborazione, indifferenza, guerra) si fossero stabilite tra i due gruppi umani. Pare indubbio, comunque, che le pulsioni artistiche furono comuni a entrambe le specie.
L'arte del Paleolitico si suole convenzionalmente dividere in due gruppi: arte parietale ed arte mobiliare. L'arte parietale è costituita da quattro periodi (detti anche stili) con una certa evoluzione temporale:
- I stile: L'arte parietale non è ancora tale, i disegni vengono realizzati su massi;
- II stile: Sorge la vera arte parietale, con graffiti sulle pareti;
- III stile: Netta evoluzione dell'arte parietale, nuove raffigurazioni di mammiferi: corna raffigurate di profilo, testa piccola, ventri enormi e zampette che sbucano dai ventri;
- IV stile: L'ultima, grande evoluzione dell'arte parietale: miglior utilizzo della prospettiva e delle tecniche di luce.
L'arte parietale era, per gli uomini del paleolitico, una rappresentazione del soprannaturale, i cui principi conduttori erano quelli dell'elemento maschile, rappresentato dal cavallo, e femminile, rappresentato dal bisonte.
Bisogna ricordare in particolare una caratteristica di alcune tecniche dell'arte parietale, il negativo, ottenuto appoggiando la mano sul muro e tutt'attorno spruzzando del colore, usando probabilmente la bocca.
L'arte mobiliare: caratterizzata dalla rappresentazione di piccole statuette, le cosiddette Veneri, forse collegate al culto della fecondità e possibile funzione religiosa oltre che ornamentale. Tra queste ricordiamo la Venere di Willendorf, che si stima sia stata realizzata fra il 23.000 e il 19.000 a.C.

Venere di Willendorf, che si stima sia stata realizzata fra il 23.000 e il 19.000 a.C.
Produzione litica del paleolitico
Il paleolitico è caratterizzato dalla realizzazione degli strumenti in pietra con la tecnica della pietra scheggiata (come il choppers: realizzato dalla prima forma di evoluzione dell'uomo: l'ominide). Questa tecnica fu ancora utilizzata nei periodi successivi, ma mescolata ad altre di più recente introduzione.
La classificazione dei manufatti può seguire le liste tipologiche dell'archeologo francese Bordes (suddivisa in strumenti su scheggia, nuclei e bifacciali) (I bifacciali erano ottenuti lavorando su due facce frammenti di pietra (schegge), ciottoli o nuclei di selce con ritocco bifacciale coprente o invadente, in modo da ottenere strumenti con margini a spigolo vivo ad andamento convesso o rettilineo, convergenti verso un'estremità. La loro forma risulta varia a seconda delle dimensioni, dello spessore e dell'andamento dei bordi e delle estremità), di Broglio-Kozlowski (suddivisa in pre-nuclei e nuclei, strumenti e armature) e di de Sonneville Bordes-Perrot.
Le tecniche di scheggiatura possono essere: "a percussione diretta", "a percussione indiretta", "a percussione su incudine", "a percussione bipolare", "a pressione".
Nel paleolitico inferiore gli utensili sono realizzati con ciottoli scheggiati (cultura dei ciottoli, o "Pebble Culture") o manufatti a forma di mandorla (bifacciali o amigdale); nel paleolitico medio con la lavorazione delle schegge staccate da un nucleo e nel paleolitico superiore con la lavorazione delle lame.
Paleolitico inferiore
Da circa 2,5 milioni di anni fa a circa 120.000 anni fa, il paleolitico corrisponde al Pleistocene inferiore e medio e alle glaciazioni di Günz, Mindel e Riss con i periodi interglaciali intermedi. In questo periodo si diffondono l'Homo habilis (Vedi sotto; è una specie di ominide estinta del genere Homo, apparsa nel Pleistocene, piano Gelasiano, e vissuta da circa 2,4 a 1,44 milioni di anni fa. L'olotipo di questo ominide è il fossile OH 7, trovato da Jonathan Leakey il 4 novembre 1960 nella Gola di Olduvai, in Tanzania) e l'Homo erectus (è una specie di ominide estinta appartenente al genere Homo. Un'ipotesi sostiene che sia la medesima specie di Homo ergaster, (vedi sotto) mentre un'altra che sia una specie prettamente asiatica evolutasi da Homo ergaster. Originariamente venne denominato pitecantropo e Uomo di Giava. La capacità cranica di Homo erectus era di poco superiore a quella di Homo ergaster, cioè dagli 813 cm³ ai 1.059 cm³, e si ritiene comunemente che sia stato il primo a lavorare e utilizzare pietre bifacciali e a usare il fuoco; queste innovazioni gli hanno permesso probabilmente la lavorazione delle pelli e un uso più elaborato degli alimenti rispetto agli uomini primitivi precedenti. Tra 1,8 e 1,3 milioni di anni fa l'Homo erectus migrò dall'Est Africa. Vi è chi sostiene che, specie autoctona asiatica, Homo erectus sia poi migrato in Africa).
Olduvaiano (Pebble Culture), 2.500.000-750.000 anni fa circa: manufatti su ciottoli appena scheggiati ("choppers" e "chopping tools"). Il nome deriva dal sito delle "gole di Olduwai" (o Olduvai, Tanzania). In Italia, sono stati ritrovati reperti risalenti a questo periodo, ad esempio nella zona di Monte Poggiolo, nel forlivese, e a Zambrone in provincia di Vibo Valentia.
Acheuleano, 750.000-120.000 anni fa circa: manufatti litici a forma di mandorla e lavorati su due lati in modo simmetrico ("bifacciali" o "amigdale") associati a diversi strumenti ricavati da schegge (raschiatoi e punte). Il nome deriva dal sito di Saint-Acheul, (presso Amiens, Francia). Geograficamente esiste una suddivisione tra "acheuleano classico" (Francia settentrionale e Inghilterra) e "acheuleano meridionale" (Francia meridionale e Spagna). Viene suddiviso cronologicamente in due principali fasi:
acheuleano "antico" o "arcaico", che tende a sostituire i termini di Abbevilliano, dal sito di Abbeville, e di Chelleano, dal sito di Chelles, entrambi in Francia):
una seconda fase più articolata, comprendente: "acheuleano medio", "evoluto" e "superiore", che continua nel paleolitico medio con l'"acheuleano finale".
Clactoniano: manufatti litici derivati da grandi schegge con piano di percussione obliquo. Secondo alcuni non si tratterebbe tuttavia di una cultura distinta dall'Acheuleano, a cui è in parte contemporaneo. Il nome deriva dal sito di Clacton-on-Sea (contea di Essex, Gran Bretagna). A volte suddiviso in "antico", "medio" e "recente".
Tayaziano: manufatti di tipo clactoniano associati ad altri di tipo musteriano, con basse percentuali di bifacciali. Non è chiaro se si tratti di una cultura autonoma e in quali rapporti sia con l'acheuleano. Il nome deriva dal sito di Les-Eyzies-de-Tayac in Dordogna, Francia.

Primi utensili in semplice pietra scheggiata, Paleolitico Inferiore
Paleolitico medio
Da circa 300.000 a circa 36.000 anni fa, corrisponde a parte del Pleistocene superiore comprendente il periodo interglaciale di Riss-Würm e parte del periodo glaciale di Würm. In questo periodo si diffonde in Europa l'Homo neanderthalensis.
Fasi finali dell'acheuleano "acheuleano finale" e "micocchiano" (130.000-70.000 anni fa circa), dal sito di La Micoque in Dordogna, Francia.
Musteriano, da circa 120.000 a circa 40-35.000 anni fa: manufatti caratterizzati da un perfezionamento delle tecniche di lavorazione (scheggiatura "levalloisiana" o "tecnica Levallois", dal sito di Levallois, differenziazione degli strumenti su scheggia, aumento degli strumenti derivati da lama). Il nome deriva dal sito di Le Moustier in Dordogna, Francia. Si suddivide in:
"musteriano di tradizione acheuleana"
"musteriano laquiniano", dal sito di La Quina, o "charentiano", dal dipartimento francese della Charente in cui si trova il sito citato.
"musteriano tipico"
"musteriano denticolato" o "a denticolati".
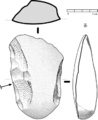
Coltello del paleolitico medio
Paleolitico superiore
Da circa 36.000 a circa 12.000 anni fa; corrisponde a parte del Pleistocene superiore comprendente parte del periodo glaciale di Würm. In questo periodo si diffonde in Europa l'odierno Homo sapiens.
Castelperroniano (40.000-34.000 anni fa circa) in Francia e Spagna nord-occidentale, dal sito di Châtelperron in Francia (considerato da alcuni "perigordiano inferiore"), e Uluzziano (38-36 000 33-30 000 anni fa circa) nell'Italia centro-meridionale, dal sito della baia e della grotta di Uluzzo, in Puglia, rappresentano culture di transizione dalle culture musteriane, ad opera ancora dei neanderthaliani e con il perdurare della tecnica levalloisiana. Un'altra cultura di transizione è lo Szeletiano (o "Szeliano", 40 000-30 000 anni fa circa) nell'Europa centrale, dal sito della grotta Széléta in Ungheria.
Aurignaziano (o "aurignaciano") (39-34 000 26-21.000 anni fa circa), con manufatti litici ricavati soprattutto da lame e microlamine e la diffusione dei manufatti in osso. Dal punto di vista geografico è suddiviso in "occidentale", "centro-europeo e balcanico", "italiano" e "orientale". Il nome deriva dal sito di Aurignac in Francia. Cronologicamente suddiviso in:
"aurignaziano arcaico" (o "pre-aurignaziano o "proto-aurignaziano")
"aurignaziano classico" ("antico", I e II, ed "evoluto", III e IV,)
"aurignaziano tardivo" (V).
Gravettiano (o "perigordiano superiore") (29-28.000 22-20.000 anni fa), caratterizzato da bulini, punte ritoccate (punte gravettiane) e armi da lancio in osso. A questa cultura appartengono molte delle più note veneri paleolitiche. Dal sito di La Gravette, presso Bayac, in Dordogna, Francia. Viene suddiviso in:
"gravettiano antico"
"gravettiano evoluto"
"gravettiano finale".
Solutreano (21-20.000-18.000 anni fa circa), caratterizzato dalla tecnica di scheggiatura a pressione, che consente di ottenere manufatti di grande raffinatezza. Viene utilizzato anche l'osso (aghi) e il corno. Compaiono i primi esempi di arte rupestre (pitture nelle caverne). Il nome deriva dal sito di Solutré, presso Mâcon, in Francia. Viene cronologicamente suddiviso in:
"proto-solutreano"
"solutreano inferiore"
"solutreano medio"
"solutreano superiore".

Arpioni magdaleniani in osso di renna a seghettatura singola e doppia.
Magdaleniano (o "maddaleniano") (18.000 11.000 anni fa, verso la fine dell'ultima glaciazione), caratterizzato dalla lavorazione di lame e nelle fasi intermedie di manufatti di piccole dimensioni ("microliti"). Si diffonde la lavorazione dell'avorio e dell'osso, con raffinata decorazione e vengono realizzate collane con denti di carnivori. A questo periodo appartiene la fioritura dell'arte rupestre (pitture nelle caverne). Il nome deriva dal sito di Abri de la Madeleine, presso Tursac, in Dordogna, Francia. In Italia è facile trovare il termine "romanelliano" che deriva dalla Grotta Romanelli in Puglia. Viene suddiviso, non unanimemente, in
"magdaleniano antico" (I-III)
"magadaleniano recente" (IV-VI)
"magdaleniano terminale", o "aziliano", dal sito di Le Mas-d'Azil nei Pirenei francesi, secondo alcuni già nel Mesolitico.
In Italia e in Europa centro-orientale mancano il solutreano e il magdaleniano: il periodo tra 20 000 e 10 000 anni fa vede una tarda evoluzione del gravettiano, l'Epigravettiano.
Viene cronologicamente suddiviso in
"epigravettiano antico"
"epigravettiano evoluto"v
"epigravettiano finale".
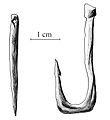
Ago e amo in osso, Paleolitico Superiore
L'uomo di Altamura
Si tratta di un esemplare umano adulto, intrappolato nella grotta di Lamalunga in Altamura, rimasto inglobato nelle stalattiti e stalagmiti che gli sono cresciute intorno e che lo hanno conservato intatto. L'equipe guidata da Vittorio Pesce Delfino dell'Università di Bari avanzò, subito dopo la scoperta, la prima stima, su basi esclusivamente morfologiche, del reperto; considerandolo una forma di pre-neandertaliano. Secondo questa ipotesi l'Uomo di Altamura doveva collocarsi antecedentemente alle forme più antiche di Neanderthal classici e successivamente alle fasi corrispondenti all' Homo erectus. Di conseguenza la stima della datazione prevedeva un intervallo tra 400.000 e 100.000 anni fa, con valori più probabili intorno a 150-250.000 anni fa. Questi primi studi, condotti preservando il reperto nel suo sito di ritrovamento, evitando in modo assoluto la rimozione di frammenti ossei o di connesse concrezioni calcaree, hanno permesso di riconoscere con certezza i tipici caratteri neanderthaliani (morfologia delle orbite e degli ispessimenti ossei sopraorbitari, assenza di fossa canina e presenza di uno spigolo ben evidente sull'osso mascellare, ispessimento dell'osso occipitale, caratteristica della apofisi mastoide, esistenza di uno spazio retromolare e andamento del margine superiore della branca ascendente della mandibola). Alcune caratteristiche del reperto associano caratteri che tipicamente ricorrono in Homo sapiens, tra i quali, in particolare, la convessità della squama dell'osso occipitale. Gli ultimi studi, eseguiti viceversa analizzando in laboratorio reperti di facile rimozione, hanno indicato una datazione meno incerta ma non in contraddizione con la prima stima del Prof Vittorio Pesce Delfino; le analisi sugli strati di calcite depositatisi attorno al reperto, effettuate nel 2015 da un team guidato da Giorgio Manzi, hanno determinato con certezza che lo scheletro, riferibile a un Neanderthal, risale a un periodo fra i 128.000 e i 187.000 anni fa.
L'interesse dei media verso il reperto paleoantropologico Uomo di Altamura deriva da numerosi fattori quali la spettacolarità naturalistica dell'intero complesso rappresentato dalle ossa nell'ambiente carsico che le ha concrezionate, saldandole le une alle altre e rendendole assolutamente fisse, la completezza dello scheletro, e le caratteristiche morfologiche richiamate.
LINEA EVOLUTIVA FINO ALL'HOMO SAPIENS
Australopitechi
L'australopiteco (Australopithecus Dart, 1925) è un genere estinto di primati della famiglia degli ominidi, che si ritiene appartenente alla linea evolutiva dell'uomo. Il nome significa "scimmia del sud". Gli australopitechi apparvero all'incirca 4,2 milioni di anni fa con l'Australopithecus anamensis (da alcuni autori ascrivibile però al genere Praeanthropus) ed ebbero un certo successo evolutivo divenendo assai diffusi in Africa, fino ad estinguersi completamente circa 2 milioni di anni fa. Le attuali conoscenze sull'origine dell'uomo permettono di tracciare, nelle sue tappe essenziali, la storia evolutiva della famiglia di primati di cui facciamo parte, quella degli ominidi, a iniziare dalle fasi molto antiche in cui comparvero gli Australopitechi, probabilmente di poco posteriori al punto di separazione dalla linea che ha condotto ai nostri parenti viventi più prossimi, gli scimpanzé.
Il primo esemplare fossile di australopiteco venne scoperto negli anni venti in un deposito calcareo a Taung, in Sudafrica, dall'antropologo australiano Raymond Dart: si trattava di un giovane di tre anni, che assieme a tratti umanoidi presentava numerose caratteristiche tipiche delle scimmie antropomorfe. Dart ipotizzò che il giovane fosse una sorta di "anello mancante" e gli assegnò il nome scientifico di Australopithecus africanus.
Nel 1935, il paleontologo scozzese Robert Broom ritrovò numerosi resti di conspecifici del cosiddetto "bambino di Taung" scoperto da Dart dieci anni prima, oltre a fossili di una nuova specie che classificò come Paranthropus (Australopithecus robustus). Per tutto il decennio successivo alla scoperta di questi ominidi, nella comunità scientifica si discusse animatamente sulla loro posizione filogenetica, in quanto molti studiosi erano eticamente contrari all'accettazione delle specie da poco scoperte come qualcosa di diverso da scimmie preistoriche.
Nel 1959, nella gola di Olduvai (Tanzania), Mary Leakey riportò alla luce un cranio di una nuova specie, battezzata Australopithecus boisei: continuando a scavare, negli anni successivi vennero rinvenuti nello stesso sito altri australopitecidi, così come esemplari di Homo habilis ed Homo erectus.
Il 24 novembre 1974 ad Hadar, nel triangolo di Afar in Etiopia, i paleontologi Yves Coppens, Donald Johanson, Maurice Taïeb e Tom Gray rinvennero i resti di un esemplare femmina dell'età apparente di 18 anni di una nuova specie vissuta circa 3,2 milioni di anni fa (Piacenziano) che fu chiamata Australopithecus afarensis. Al fossile venne dato il nome di Lucy, in onore della canzone Lucy in the Sky with Diamonds dei Beatles, mentre in amarico è noto come Dinqinesh, che significa "Tu sei meravigliosa". Il nome in codice è A.L. 288 (Afar Locality n° 288). I resti comprendevano circa il 40% dello scheletro (52 ossa); particolarmente importanti l'osso pelvico, il femore e la tibia, perché la loro forma lascia pensare che questa specie fosse già bipede.
Successivamente i ricercatori hanno scoperto altre specie di australopitecidi, che hanno contribuito a fare maggiore chiarezza sull'esatto periodo durante il quale i membri di questo genere sono vissuti: ad esempio, il ritrovamento dei resti di Australopithecus sediba, risalenti a 1,9 milioni di anni fa (si pensava che gli australopitecidi si fossero estinti prima di 2 milioni di anni fa) in Sudafrica ha dimostrato che questi ominidi sono vissuti per molto tempo dopo la loro presunta scomparsa, convivendo fra loro e anche con alcune specie del genere Homo.
Ipotesi evolutiva degli australopitecini secondo il paleoantropologo tedesco Friedemann Schrenk, dove H. habilis viene ascritto ad un ramo evolutivo strettamente australopiteco. La scala temporale a destra è in milioni di anni.
Il nome del genere deriva dalla combinazione delle parole australis, che in latino significa " nativo dell'emisfero meridionale", e πίθηκος (pithekos), che in greco significa "scimmia": Australopithecus significa pertanto "scimmia australe", in riferimento al fatto che i primi resti fossili di specie ascrivibili al genere finora rinvenuti sono stati trovati nella porzione australe del continente africano.
Al genere vengono ascritte nove specie, tutte estinte, raggruppate in due suddivisioni, valutate oggi generi distinti:
1. Australopithecus
Australopithecus afarensis
Australopithecus africanus
Bambino di Taung
Australopithecus anamensis
Australopithecus bahrelghazali
Australopithecus garhi
Australopithecus sediba
2. Paranthropus
Australopithecus aethiopicus ora Paranthropus aethiopicus
Australopithecus boisei ora Paranthropus boisei
Australopithecus robustus ora Paranthropus robustus
Di queste, appunto solo tre (A. africanus, garhi e sediba) vengono considerate come effettivamente ascrivibili al genere: alcuni studiosi ritengono maggiormente corretto ascrivere le rimanenti specie ai generi Paranthropus (A. aethiopicus, boisei e robustus) e Praeanthropus (A. afarensis, anamensis e bahrelghazali). Tale scelta sarebbe guidata da differenze abbastanza consistenti a livello morfologico e probabilmente anche ecologico fra i vari gruppi di specie, differenze che tuttavia potrebbero essere giustificabili anche considerando il vasto areale occupato dal genere e il vasto lasso di tempo durante il quale esso si è potuto evolvere e diversificare.
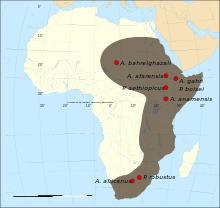
I punti rossi rappresentano i luoghi di ritrovamento delle varie specie di Australopithecus, mentre la probabile area di diffusione del genere è colorata in grigio scuro
I primi membri del genere Australopithecus si evolsero in Africa centro-orientale circa 4 milioni di anni fa. Si trattava di esseri con numerosi tratti comuni alle scimmie antropomorfe e all'uomo, con andatura fondamentalmente bipede (come intuibile dalle numerose impronte fossili scoperte nel continente africano, fra le quali particolarmente famose e ben conservate sono quelle di Laetoli, in Tanzania), ma pronti ad arrampicarsi sui radi alberi della savana per sfuggire ai predatori o per trovare un rifugio sicuro dove passare la notte.
Nonostante la taglia contenuta e la mancanza di particolari adattamenti che ne assicurassero la competitività, gli australopitecidi riuscirono ad affermarsi grazie alla dieta onnivora, che consentiva loro di trovare nutrimento in qualsiasi frangente, sfruttando indifferentemente risorse di origine animale (ad esempio carcasse di grossi erbivori uccisi dai predatori, oppure piccole prede catturate occasionalmente come roditori e uccelli ma anche bruchi e uova) così come le risorse offerte dalla terra (radici, frutti ed altri cibi di origine vegetale). Questo opportunismo permise agli australopitecidi di diffondersi in gran parte del continente africano.
Gli studiosi sono propensi a credere che dal genere Australopithecus, e in particolare dalla specie africanus, si siano staccati i progenitori del genere Homo, attorno ai due milioni di anni fa; ciò è verosimile, tuttavia sono stati rinvenuti resti fossili di primati ascrivibili al genere Homo, ma antecedenti all'apparizione di Australopithecus africanus. Questo indicherebbe che il distacco dagli australopitecini degli antenati dell'uomo moderno potrebbe essere avvenuto prima di quanto si pensasse, ad esempio a partire da Australopithecus afarensis, o da specie ancora più primitive addirittura estranee al genere, come Kenyanthropus platyops. Anche l'apparizione degli australopitechi del ramo Paranthropus può essere vista come un distacco dalla linea originaria, in virtù del netto cambiamento nelle abitudini e nella morfologia che contraddistingue le specie di questo genere da quelle del ramo ancestrale.
Morfologia e comportamento
Si trattava di animali piuttosto piccoli e gracili, di altezza compresa fra i 120 e i 150 cm. Era presente un dimorfismo sessuale piuttosto accentuato, coi maschi considerevolmente più grandi e robusti delle femmine (fino al 50%, contro una media del 15% nell'uomo moderno). Ciò lascia supporre che questi animali vivessero in gruppi capitanati da un maschio dominante, similmente a quanto osservabile fra gli attuali gorilla.
Il cervello della maggior parte degli Australopitecus aveva dimensioni pari a circa il 35% di quelle dell'attuale cervello del genere Homo. La mandibola era molto robusta e munita di denti forti ed appiattiti, con canini poco pronunciati e premolari e molari forti e dallo smalto ispessito, indicanti una dieta principalmente vegetariana. Gli arti anteriori avevano pressappoco la stessa lunghezza di quelli posteriori, nei quali l'opponibilità del pollice era stata praticamente persa per supportare un'andatura bipede.
Non si ha notizia di utensili utilizzati dagli australopitechi (o almeno si pensa che essi non fossero maggiormente dediti al loro utilizzo di quanto non lo siano i primati moderni), così come si pensa che essi non abbiano sviluppato alcuna forma di linguaggio. Australopithecus garhi sembrerebbe tuttavia essere un'eccezione: i resti di questa specie sono stati ritrovati assieme a utensili e resti di animali macellati, il che farebbe pensare al sorgere di una primitiva industria degli utensili sviluppata parallelamente a quella di Homo, in quanto A. garhi si sarebbe evoluto molto probabilmente dopo il distacco dagli australopitechi della linea evolutiva che avrebbe poi portato a Homo.
Il fatto che gli australopitechi fossero fondamentalmente degli scimpanzé bipedi significa che l'evoluzione di un'andatura bipede non è stata influenzata in modo significativo dall'aumento in capacità della scatola cranica e quindi dall'accrescimento dell'intelligenza, come veniva invece propugnato fino a tempi recenti da numerosi studiosi. Tale ipotesi era stata fra l'altro già messa in discussione dal ritrovamento di Orrorin tugenensis, primate bipede vissuto circa 6 milioni di anni fa.
La spiegazione più accreditata sull'acquisizione di un'andatura bipede indica questa caratteristica come un adattamento all'avanzata della savana in seguito ai cambiamenti climatici che interessarono l'Africa centro-orientale attorno ai 10 milioni di anni fa: l'andatura eretta consentiva agli australopitechi di ergersi al di sopra dell'erba alta e osservare agevolmente i dintorni, individuando fonti di cibo o di pericolo. Alcuni studiosi hanno però osservato che per gli ominidi primitivi sarebbe stato molto difficile, se non impossibile, sostenere un cambiamento così veloce nel tempo (in termini evolutivi) sia a livello morfologico (acquisizione di un'andatura bipede, con annessi cambiamenti a livello osteo-muscolare) che a livello comportamentale (migrazione dalla foresta pluviale alla savana semiarida). Si pensa perciò che l'andatura bipede fosse già in fase di acquisizione quando la savana lambì le aree dove vivevano i progenitori degli australopitechi: osservando due specie di scimmie antropomorfe attuali, come l'orango e lo scimpanzé, si nota infatti che il primo tende letteralmente a camminare orizzontalmente fra i rami, muovendosi su di essi con le sole zampe posteriori e mantenendosi alle liane ed ai rami verticali con le braccia, mentre il secondo si arrampica verticalmente abbracciando il ramo e puntellandosi con le zampe posteriori. Si può quindi pensare che nei primi australopitechi la forte muscolatura delle gambe fosse evoluta come adattamento al movimento orizzontale sui rami della volta arborea (attività che non richiede certo un'intelligenza elevata) e che in un secondo momento essa sia tornata assai utile per muoversi al suolo nelle sterminate pianure africane.
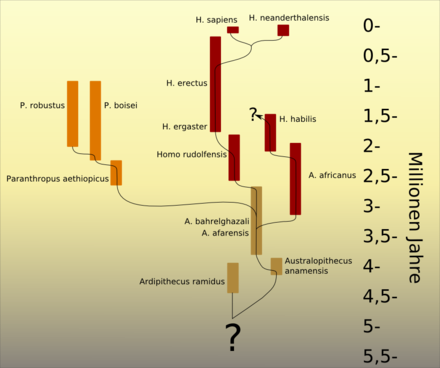
Ipotesi evolutiva degli australopitecini secondo il paleoantropologo tedesco Friedemann Schrenk, dove Homo habilis viene ascritto ad un ramo evolutivo strettamente australopiteco. La scala temporale a destra è in milioni di anni
Homo habilis
Homo habilis è una specie di ominide estinta del genere Homo, apparsa nel Pleistocene, piano Gelasiano, e vissuta da circa 2,4 a 1,44 milioni di anni fa.
L'olotipo di questo ominide è il fossile OH 7, trovato da Jonathan Leakey il 4 novembre 1960 nella Gola di Olduvai, in Tanzania. Dallo studio dei fossili è stato subito evidente come Homo habilis presenti un insieme di caratteri plesiomorfi e derivati. Va notato che i caratteri derivati, ovvero nuovi rispetto agli australopitechi, si concentrano principalmente nella morfologia cranica. Infatti in questa specie il volume encefalico medio è di 612 cm³ che, sebbene inferiore alla media degli uomini moderni, è decisamente superiore a quella delle austrolopitecine, che, a esclusione di P. boisei, non superavano la soglia dei 500 cm³, considerata da alcuni paleoantropologi uno dei caratteri discriminanti per l'attribuzione di una specie al genere Homo. Inoltre la volta cranica risulta essere più arrotondata che nel genere Australopithecus, anche se la massima espansione è ancora molto bassa, e permane una moderata costrizione retrorbitale. La cresta sagittale, punto di attacco dei muscoli temporali, è assente. Il toro sopraorbitario è presente anche se decisamente ridotto rispetto alle australopitecine. Dietro al toro è presente il solco retrotorale dal quale parte la fronte che ha una crescita più verticale delle specie precedenti sebbene essa sia ancora sfuggente se comparata alle specie successive. La faccia presenta dimensioni ridotte rispetto alle australopitecine così come presenta un minore prognatismo mediofacciale. La fossa canina e il mento, carattere tipico di Homo sapiens, sono assenti. Homo habilis presenta una arcata dentale pressoché parabolica con denti che presentano una ridotta megadonzia e in generale sono più moderni che nelle specie di ominidi fino a quel momento vissute, infatti presenta incisivi larghi e molari e premolari allungati e stretti.
Il record fossile non ci ha restituito molto del post-craniale e la maggior parte delle informazioni al riguardo che abbiamo provengono dal fossile OH 62 (conosciuto come Lucy's Child), ritrovato nella Gola di Olduvai in Tanzania da Tim White durante la campagna di scavi del 1986. Lucy's Child è composto da più di 300 frammenti tra i quali sono presenti l'omero, il radio e l'ulna dell'arto destro, quasi completi e la parte prossimale del femore sinistro. Dalla circonferenza del femore si è potuto stabilire che l'altezza di OH 62, e presumibilmente degli altri Homo habilis, fosse intorno ai 100-135 cm. Le ossa dell'arto superiore risultano molto lunghe, soprattutto se paragonate all'arto inferiore; infatti il rapporto tra omero e femore si attesta intorno allo 0,95 contro lo 0,75 degli uomini attuali, carattere quindi decisamente australopitecino. Le informazioni sulla forma del piede, carattere importante per decifrare il tipo di andatura, provengono quasi unicamente dal fossile OH 8. Esso mostra una struttura moderna ed è quindi possibile asserire con certezza quasi assoluta che Homo habilis avesse una andatura bipede, anche se molto probabilmente non perfetta.
Dotato di una capacità cranica di circa 600750 cm³, Homo habilis utilizzava i suoi strumenti per uccidere e squartare le carcasse di animali. Tali manufatti erano ancora abbastanza primitivi, ma il fatto che tali ominidi li costruissero implica delle importanti considerazioni:
- H. habilis prefigurava la necessità futura di tali oggetti;
- H. habilis sapeva scegliere i materiali disponibili per costruirli;
- H. habilis possedeva l'abilità manuale e cognitiva per realizzarli secondo necessità.
- Si ritiene che Homo habilis fosse in grado di padroneggiare gli utensili di pietra del primo Paleolitico. Si trattava degli utensili più avanzati mai usati, e diedero a habilis la capacità di prosperare in un ambiente ostile, in precedenza troppo pericoloso per i primati. È ancora controverso se habilis sia stato il primo ominide in grado di padroneggiare gli utensili di pietra, poiché fossili di Australopithecus garhi, datati 2,5 milioni di anni fa, sono stati ritrovati con frammenti di utensili in reperti più antichi di almeno 100.000-200.000 anni di Homo habilis.
La maggior parte degli esperti sostiene che l'intelligenza e l'organizzazione sociale di Homo habilis fosse più sofisticata di quella degli australopitecidi e degli scimpanzé. Eppure, nonostante l'utilizzo di strumenti, Homo habilis non era un abile cacciatore rispetto ad altre specie. Sembrerebbe che utilizzasse gli utensili soprattutto per strappare la carne della preda, piuttosto che per difesa o per cacciare. Homo habilis era forse l'antenato del più avanzato Homo ergaster, che a sua volta fu l'antenato dell'Homo erectus. Continuano ad esserci dibattiti sulla tesi che habilis sia stato o meno un diretto antenato dell'uomo, e vi sono anche dubbi che tutti i fossili noti siano stati attribuiti correttamente a questa specie.
Homo habilis coesisteva con altri bipedi primati - come il Paranthropus boisei - alcuni dei quali prosperarono per millenni. Tuttavia habilis, forse a causa dell'utilizzo di utensili e per la sua dieta meno specializzata, divenne il precursore di un'intera linea di nuove specie, mentre il Paranthropus boisei e le specie correlate scomparvero.
La classificazione di Homo. habilis nel genere Homo è tuttora controversa: alcuni paleontologi preferiscono parlare di Australophitecus habilis. Come Homo rudolfensis, omo. habilis mancava di molte caratteristiche tipiche degli ominidi più evoluti, come le ossa delle anche più strette per camminare meglio per lunghe distanze, un sistema di sudorazione più sofisticato, un canale del parto più stretto e gambe più lunghe delle braccia.
Fossili
I fossili di Homo habilis sono stati ritrovati in varie parti dell'Africa orientale e meridionale; di particolare rilevanza sono i siti della Chemeron Formation, presso Tugen Hill e dell'area del Lago Turkana (già Lago Rodolfo) in Kenya; della valle dell'Omo e Middle Awash in Etiopia; dell'Uraha in Malawi; di Sterkfontein in Sudafrica e della Gola di Olduvai in Tanzania, da cui Mary Leakey e la sua squadra recuperarono i primissimi fossili di questa specie. ]
Di seguito sono elencati alcuni dei principali fossili di H. habilis:
- OH 7 (Jonny's Child): olotipo della specie, è stato scoperto da Jonathan Leakey il 4 novembre 1960 nella Gola di Olduvai in Tanzania. È composto da un osso parietale destro quasi completo, un parietale sinistro molto frammentato, la mandibola quasi completa, tutti i denti inferiori e alcuni denti superiori (incisivi destri, canino destro, premolari destri, primo molare destro e secondo molare sinistro). OH 7 è datato 1.75 Ma.
- OH 8: scoperto dal gruppo di ricerca di Leakey nel 1960 nella Gola di Olduvai, in Tanzania, a 5 metri dalla mandibola di OH 7. Per questo alcuni studiosi ritengono che i due fossili appartengano allo stesso individuo. Datato 1.75 Ma il fossile è un piede sinistro parzialmente integro di un individuo giovane. OH 8 è composta dall'astragalo, dall'osso navicolare, dal calcagno, dall'osso cuboide, dalle ossa cuneiformi (mediale, intermedio e laterale) e dalle cinque ossa metatarsali.
- OH 24 (Twiggy): Cranio fossile frammentato datato 1,8 Ma, ritrovato da Peter Nzube nell'ottobre del 1963 nella Gola di Olduvai, in Tanzania. Al momento del ritrovamento, all'interno di un blocco di calcare, il fossile appariva schiacciato e per questo fu soprannominato Twiggy, come la magrissima modella londinese Twiggy Lawson.
KNM-ER 1813: scoperto da Kamoya Kimeu nel 1973 a Koobi Fora, nei pressi del lago Turkana, in Kenya. Datato 1,65 Ma, questo fossile è composto da un cranio di adulto quasi completamente conservato e i danni maggiori riguardano la parte sinistra della faccia. Il volume encefalico è di 510 cm³, ovvero al disotto della media della specie. Il fossile è conservato al Museo nazionale del Kenya (KNM).
KNM-ER 1805: cranio di un individuo adulto diviso in tre frammenti principali (cranio, mandibola e mascella). Ritrovato nel 1973 a Koobi Fora, nei pressi del lago Turkana in Tanzania, da Paul Abell ed è datato 1,7 Ma.
Homo ergaster
L'Homo ergaster visse probabilmente tra i due milioni di anni ed un milione di anni. Si stabilì in molte zone del continente africano, comprese tra l'Africa orientale ed il Sudafrica (Arsuaga, 2001). Forse condivise alcuni di questi luoghi con altre specie, come l'Homo habilis, che 1,8 milioni di anni fa era ancora presente presso la Gola di Olduvai. La sua corporatura, dimensioni e proporzioni, era simile alla nostra, mentre la distanza dagli australopiteci e gli altri homo era abbastanza marcata. "Turkana boy" (KNM-WT 15000), lo scheletro di un bambino di 10 anni è la prova più importante. La corporatura di questo bambino corrispondeva a quella di un ragazzo moderno più grande di 1 o 2 anni. Il volume encefalico dell'Homo ergaster era maggiore che negli altri ominidi, che in alcuni casi meglio conservati è: 804 cc, 850 cc e 900 cc (Arsuaga, 2001). In termini relativi questo risultato va ridimensionato. Il cervello dell'Homo ergaster cresce in proporzione al corpo, quindi non si verifica nessun progresso significativo rispetto all'Homo habilis. Tuttavia, si verificò un notevole balzo in avanti delle capacità cognitive. Secondo alcuni questo cambiamento fu maggiore nei maschi che nelle femmine, soprattutto riguardo al senso dell'orientamento, alla capacità di ricordare luoghi o la posizione degli oggetti.
I resti fossili più importanti di Homo ergaster sono principalmente due. Tutti e due sono stati scoperti in Kenia, tra il 1975 e il 1984. Il secondo, scoperto nei pressi del Lago Turkana, è composto da uno scheletro di un bambino di circa dieci anni, (KNM-ER 3733 e KNM-WT 15000). Tali resti risalgono ad un periodo compreso tra gli 1,8 e i 1,6 milioni di anni fa, mentre altri fossili dello stesso periodo sono stati attribuiti ad Homo erectus (OH 9 e OH 12).
- KNM-ER 3733. Scoperto da Bernard Ngeneo nel 1975 a Koobi Fora in Kenia. L'eta stimata è di 1.7 milioni di anni. La scoperta straordinaria consiste in un cranio molto completo. Il volume del cranio è di 850 cc., e il teschio completo è molto simile all'uomo di Pechino. La scoperta di questo fossile nello stesso strato di ER 406 (Australopithecus boisei) diede il colpo di grazia all'Ipotesi dell'unica Specie: L'idea che ci poteva essere un'unica specie di ominidi in ogni punto della storia.
- KNM-WT 15000, detto "Turkana Boy". Scoperto da Kamoya Kimeu (della èquipe di Richard Leakey) nel 1984 a Nariokotome vicino al Lago Turkana in Kenia. Questo è uno scheletro completo di un bambino di 11 o 12 anni, l'unica grande omissione sono le mani e i piedi. (Molti scienziati pensano che gli Erectus maturano più in fretta degli uomini moderni, ed egli aveva in realtà soltanto 9-10 anni). È il più completo scheletro di ergaster conosciuto, ed è anche uno dei più vecchi, 1.6 milioni di anni. Il volume del cranio era di 880 cc, ed è stato stimato che poteva diventare 910 cc da adulto. Il ragazzo era alto 1,60 m e sarebbe diventato 1,85 m da adulto. Questa è un'altezza sorprendente, indica che molti Erectus potevano essere più grossi degli uomini moderni. Ad eccezione del teschio, lo scheletro era molto simile ad un ragazzo moderno, sebbene potesse avere delle piccole differenze.
L'Homo ergaster sembra essere stato carnivoro a differenza degli ominidi.
Homo erectus
Homo erectus è una specie di ominide estinta appartenente al genere Homo. Un'ipotesi sostiene che sia la medesima specie di Homo ergaster, mentre un'altra che sia una specie prettamente asiatica evolutasi da ergaster. Originariamente venne denominato pitecantropo e Uomo di Giava. La capacità cranica di erectus era di poco superiore a quella di ergaster, cioè dagli 813 cm³ ai 1.059 cm³, e si ritiene comunemente che sia stato il primo a lavorare e utilizzare pietre bifacciali e a usare il fuoco; queste innovazioni gli hanno permesso probabilmente la lavorazione delle pelli e un uso più elaborato degli alimenti rispetto agli uomini primitivi precedenti. Tra 1,8 e 1,3 milioni di anni fa Homo erectus migrò dall'Est Africa. Vi è chi sostiene che, specie autoctona asiatica, erectus sia poi migrato in Africa. La sua scoperta risale al 1891, quando, nel giacimento di Trinil dell'isola di Giava, Eugène Dubois rinvenne una calotta cranica, insieme a un molare e un femore. Dalle conoscenze fino ad allora accumulate egli desunse che si trattasse di un uomo scimmia, per cui gli diede il nome di Pithecanthropus erectus. Oggi noi sappiamo tuttavia che Homo erectus, come è stato poi ribattezzato, era un ominide più evoluto rispetto al genere Australopithecus.
Altri ritrovamenti:
OH 9, Chellean men. Scoperto da Louis Leakey nel 1960 nella Gola di Olduvai in Tanzania. L'età stimata è di 1,4 milioni di anni. Esso consiste in parte della scatola cranica con grosse arcate frontali e un cervello di 1065 cm³.
OH 12, Pinhead. Scoperto da Margaret Cropper nel 1962 nella Gola di Olduvai in Tanzania. Esso è simile a OH 9 ma è più completo e piccolo, con una scatola cranica di soli 750 cm³. La sua età è compresa tra i 600 000 e gli 800 000 anni.
Nel 2000, nella Cina meridionale, sono emersi una serie di fossili litici non bifacciali datati tra i 700.000 e gli 800.000 anni fa, che i due scopritori, Huang Weiwen e Rick Potts, propongono di assegnare al modo tecnico acheuleano. Tra le cause di questa importante differenza vi può essere un impedimento oggettivo, come la mancanza di materiale utile per la costruzione di questi attrezzi o l'impossibilità della trasmissione di questa conoscenza da una generazione all'altra. Un'altra tesi è che la colonizzazione dell'Asia sia antecedente alla scoperta delle asce a mano avvenuta in Africa e che i colonizzatori siano rimasti isolati dai loro cugini africani. Negli ultimi anni sono stati fatti degli importanti ritrovamenti che confermano tale ipotesi, anticipando di alcune centinaia di migliaia di anni la colonizzazione dell'Asia. Il più importante è un teschio ritrovato nel 2001 a Dmanisi in Georgia, risalente a 1,8 milioni di anni fa e attribuito alla specie Homo georgicus. Con un volume di 600 cm³ è il fossile più antico ritrovato fuori dall'Africa; i suoi tratti somatici sembrano essere comuni a quelli degli Homo ergaster africani. Altri fossili sono stati trovati in Cina e a Giava, alcuni dei quali molto antichi, come un cranio infantile senza faccia, risalente a 1,8 milioni di anni e alcuni resti incompleti e deformati, provenienti dall'area di Sangiran, datati 1,6 milioni di anni.
Descrizione
Homo erectus aveva una capacità cranica maggiore rispetto a Homo habilis e una notevole somiglianza con gli esseri umani moderni, ma aveva un cervello di dimensioni corrispondenti a circa il 75% di quello di Homo sapiens. I membri della specie erectus erano piuttosto alti. Il dimorfismo sessuale era leggermente più marcato che in Homosapiens, poiché i maschi erano di maggiori dimensioni rispetto alle femmine. La scoperta dello scheletro KNM-WT 15000 vicino al lago Turkana in Kenya da Richard Leakey nel 1984 fu decisiva per la conoscenza della specie.
Homo erectus usava utensili più diversificati e avanzati dei suoi predecessori. Si trattava di strumenti di pietra, un'innovazione significativa fu l'utilizzo di asce a doppio filo e pietre bifacciali scheggiate su due lati, chiamate comunemente amigdale per la loro forma a mandorla, usate come strumenti per scavare radici, tritare vegetali e probabilmente per tagliare pelli.
Un sito chiamato Terra Amata, sulla Riviera Francese, era forse occupato da membri della specie erectus, e contiene alcune tra le prime (per quanto controverse) tracce dell'utilizzo di un fuoco controllato dall'uomo. Analoghe tracce sono state rinvenute in Israele. Ciononostante, alcuni studiosi continuano a ritenere che l'utilizzo del fuoco fosse raro nella specie, e che sia stato più caratteristico di specie avanzate del genere Homo - come Homo neanderthalensis.
Inoltre, è stata avanzata l'ipotesi che erectus sia stato il primo ominide in grado di utilizzare zattere per attraversare oceani, ma la tesi rimane controversa.
Alcuni antropologi, come il già citato Leakey, credono che erectus fosse significativamente più vicino dal punto di vista della vita sociale agli uomini moderni rispetto ad altre specie precedenti. Va precisato, però, che i fossili dimostrano come l'anatomia delle vie aeree dell'erectus non gli permettesse di produrre suoni di una complessità paragonabile a quella del linguaggio moderno.
Homo neanderthalensis
Homo neanderthalensis, King, 1864, comunemente detto Uomo di Neanderthal, è un ominide strettamente affine all'Homo sapiens che visse nel periodo paleolitico medio, compreso tra i 200.000 e i 40.000 anni fa.
Prende il nome dalla valle di Neander (Neandertal) presso Düsseldorf in Germania, dove vennero ritrovati i primi resti fossili. Fu un "Homo" molto evoluto, in possesso di tecnologie litiche elevate e dal comportamento sociale piuttosto avanzato, al pari dei sapiens di diversi periodi paleolitici.
Convissuto nell'ultimo periodo della sua esistenza con lo stesso Homo sapiens, l'Homo neanderthalensis scomparve in un tempo relativamente breve, evento che costituisce un enigma scientifico oggi attivamente studiato.
I resti che diedero il nome alla specie furono scoperti nell'agosto 1856 da scavatori di calcare in una grotta denominata "Kleine Feldhofer" nella valle di Neander, nei pressi della località di Feldhof in Germania. I reperti furono mostrati a Johann Fuhlrott, insegnante di scienze, che riconobbe trattarsi di una nuova specie del genere Homo. Della scoperta dei fossili venne dato annuncio ufficiale solo il 4 febbraio 1857.
Documentata fra 130.000 (per le forme arcaiche) e 30.000 (documentata con reperti fossili) -22.000 (in assenza di fossili ma con discusse prove culturali) anni fa principalmente in Europa e Asia, e limitatamente in Africa.
Alcuni studi del 2010 suggeriscono, tra alcune ipotesi probabili relative alla vicinanza genetica tra neanderthalensis e sapiens, che ibridazioni fra i due possano avere avuto luogo nel Vicino Oriente all'incirca tra 80.000 e 50.000 anni fa, per la presenza nell'uomo contemporaneo di una percentuale compresa tra l'1 e il 4% di materiale genetico specificamente neandertaliano. Tali tracce genetiche sono presenti negli eurasiatici e nei nativi americani, ma non negli africani subsahariani: ciò suggerisce, tra almeno quattro diverse ipotesi, che l'ibridazione possa avere avuto luogo nei primi stadi della migrazione della specie umana fuori dall'Africa, presumibilmente quando venne a contatto con i Neanderthal che vivevano nel Vicino Oriente, circa 80 000 anni or sono.
In passato, la specie era stata chiamata anche "uomo di Neanderthal", dall'originale nome specifico scientifico, e Homo sapiens neanderthalensis quando era ancora considerato sottospecie dell'Homo sapiens; queste denominazioni talora si riscontrano ancora oggi e così pure avviene in altre lingue. Il problema del nome come sottospecifico non è, al contrario del primo, meramente formale, ma riflette, come verrà esposto più avanti, diversi possibili cammini evolutivi e differenti gradi di reincrocio con i sapiens.
Sulla estinzione del neanderthalensis sono in elaborazione numerose teorie.
Sintesi dell'aspetto esteriore
L'aspetto fisico esteriore del neandertaliano classico, alla luce delle conoscenze attuali, notevolmente incrementatesi rispetto alle prime ipotesi ed estrapolazioni otto-novecentesche, è quello di un uomo di altezza medio-bassa (1,60 m) perfettamente eretto e muscolarmente molto robusto, con un rapporto superficie/volume inferiore a quello delle altre specie di uomini, il che comporta una minore dispersione di calore e quindi una maggiore resistenza alle basse temperature. La statura media, in base a quarantacinque ossa lunghe di almeno quattordici maschi e sette femmine, era di circa 164168 cm per i maschi e circa 152156 cm per le femmine. I campioni di ventisei esemplari studiati nel 2010 mostrano un peso medio di 77,6 kg per i maschi e 66,4 kg per le femmine.
La testa, rispetto ad un sapiens, risulta allungata antero-posteriormente, anche se si sovrappone in genere alla variabilità sapiens, ha un volume cerebrale di 1500 cm³ in media, del 10% superiore agli uomini attuali, e arcate sopraccigliari sporgenti. Ha uno spiccato prognatismo mascellare e il mento può essere sfuggente, per lo meno nei tipi arcaici. Con il passare del tempo, in alcune zone e verso la fine del Paleolitico, si diffonde un tipo più gracile e con un mento osseo più pronunciato, mentre gli zigomi sono molto meno accentuati e le arcate sopraccigliari al contrario più sporgenti.
Una tesi esposta nel 2006 e confermata nel 2007, basata su ricerche avanzate con tecniche di biologia molecolare, ipotizza che la specie, in Europa, abbia sviluppato individui di carnagione bianca con capelli rossi: il tipo di pigmentazione è in accordo con la scarsa irradiazione solare (ultravioletta) del territorio colonizzato, analogamente alla distribuzione geografica attuale della pigmentazione nei tipi umani. Ciononostante, si è evidenziato come la variabilità genetica della popolazione neandertaliana suggerisca una variabilità del fenotipo piuttosto ampia, analogamente a quella attuale di Homo sapiens.
Recenti studi, basati sull'analisi di alcune sequenze geniche di mtDNA, suggeriscono che, senza arrivare a parlare di sottospecie, vi fu sicuramente una suddivisione in tre (o forse quattro, ma il metodo non riesce ancora a chiarire quest'ipotesi) diversi grandi gruppi di popolazioni. La reale esistenza dei gruppi sud-europeo (sud-iberico, subalpino, balcanico), centro-est europeo (dalla zona nord-iberica fino al mar Caspio) e medio-asiatico (fino ai confini orientali kazaki) in precedenza era stata frequentemente messa in discussione sulla base dei soli reperti fossili.

Ricostruzione moderna di una donna Neandertal
Tecnica, cultura e arte
L'uomo di Neanderthal inizia a evolvere in un contesto culturale acheuleano superiore, dove i manufatti bifacciali cambiano forma, migliorano la punta e diminuiscono di spessore.
Nell'industria litica compare la nuova tecnica di scheggiatura levalloisiana (da Levallois, alla periferia di Parigi). Da un nucleo litico iniziale, sgrossato fino a portarlo a una forma biconvessa, lateralmente su una faccia si staccano parallelamente a un piano di base schegge di forma regolare. Questa tecnica evolve e le forme chiamate amigdale (a mo' di mandorla) dell'Acheuleano scompaiono, anche se a sud del Sahara continuerà fino al 50. 000 a.C. circa.
In Europa, territorio principale del Neanderthal, si parla di cultura musteriana, da ritrovamenti a Le Moustier, in Dordogna. Abbiamo punte triangolari, raschiatoi (per la preparazione delle pelli) molto rifiniti, dal bordo tagliente finemente ritoccato. Il Musteriano si articola in diverse culture, geografiche e cronologiche (Musteriano di tradizione acheuleana, Musteriano tipico, Musteriano denticolato, Charentiano, Musteriano pontiniano nel Lazio eccetera).
Pare accertato, con qualche residua incertezza, il passaggio successivo al Castelperroniano, con reperti affidabilmente attribuiti ai Neandertaliani come lamette litiche, manufatti in osso e ornamenti per il corpo.
Evoluzione culturale
Sull'evoluzione culturale di Homo neanderthalensis non vi è ancora una visione condivisa. A fronte di antropologi come Ian Tattersall, che non riconoscono il raggiungimento di livelli culturali che sconfinino dalla mera tecnologia e, in particolare, non condividendo il fatto che sia stata raggiunta la visione simbolica, intesa come dimensione simbolica nel senso psicologico, premessa a riti, arte, e comportamenti relativi, vi sono invece indizi di comportamenti culturali avanzati. Le culture litiche che poi evolveranno (Castelperroniano, Aurignaziano e molto dubbiosamente Gravettiano, condivise sicuramente dai sapiens) sono quindi tuttora allo studio per la sicura eventuale attribuzione ai Neandertal; pare, con buona significatività, accertato il primo passaggio.
In sintesi ed a grandi linee, si può dire che la cultura neandertaliana dominante fu il Musteriano e che il limite convenzionale (attuale) superiore si situa fra il Castelperroniano e l'Aurignaziano. Molto diffuso era l'utilizzo delle pelli, anche per la costruzione di ripari estivi all'aperto, contrapponendosi alla pratica troglodita invernale. Si ritrovano strutture di pietre o di ossa atte ad assicurare i bordi delle pelli al suolo. Abbondanti tracce di ocra rossa fanno pensare a usi rituali e religiosi. Anche in tale ottica, si evidenzia l'inumazione come pratica diffusa, in fosse di forma ovale, con corredi funerari (cibo, corna e strumenti litici), spesso ricoperte da lastroni per sottrarre i corpi alle fiere, deposizioni di fiori (studi sui pollini in ritrovamenti in Asia Minore). Il fuoco, in cerchi di contenimento di pietre, è largamente utilizzato.
Secondo una ricerca in corso nel sito della grotta di Bruniquel in Francia, i neanderthalensis avrebbero realizzato, all'incirca 175.000 anni or sono, strutture complesse (la cui funzione è ancora oggetto di indagine) utilizzando stalagmiti appositamente spezzate e poi disposte in formazioni concentriche, dimostrando un'avanzata capacità organizzativa, costruttiva e, forse, simbolica.
Il flauto di Divje Babe I
Forse (gli studi sono ancora in corso), con i neanderthalensis si ha il primo esempio di strumento musicale non a percussione, ma intonato (in dettaglio, con quattro note compatibili, con la naturale scala diatonica greca), grazie al ritrovamento del cosiddetto flauto di Divje Babe (in Slovenia): un frammento di femore di orso delle caverne perforato regolarmente. Inizia anche l'arte figurativa in senso stretto, considerata prerogativa di sapiens, ma dalla stratigrafia recentemente attribuita anche ai neanderthalensis.

Il flauto di Divje Babe I
I recenti progressi molecolari nello studio delle popolazioni neandertaliane e la loro localizzazione geografica, uniti a quelli sull'industria litica e degli altri manufatti, permetteranno in futuro di chiarire meglio i rapporti tra le diverse culture e la loro evoluzione nello spazio e nel tempo.
Nel complesso, la tecnologia dell'Homo Neanderthalensis può riassumersi in:
- Asce a mano o amigdale: sono il resto di grossi noduli di selce, scheggiati ai bordi per ricavarne schegge più piccole.
- Punte di selce: da usare immanicate su pesanti bastoni usati come lance nella caccia a grossi animali.
- Denticolati, cioè schegge di selce senza punta col margine dentellato: sarebbero primitive seghe a mano, usate per lavorare legno, ossa e tendini.
- Raschiatoi: sono dei coltelli di selce da usare senza manico, per tagliare la carne.
- Flauti, cioè ossa lunghe forate, che sarebbero una sorta di accendini: nei fori venivano sfregati bastoncini di legno per accendere paglia e altro materiale infiammabile.
Un articolo pubblicato su Proceedings of the National Academy of Sciences il 13 novembre 2012 a cura di un gruppo del Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology di Lipsia, getta nuova luce sulla sempre più probabile attività simbolica e artistica dei Neandertal nel passaggio dal Paleolitico medio (MP) a Paleolitico superiore (UP), periodo caratterizzato dalla sostituzione degli ultimi Neandertal con gli esseri umani moderni in Europa tra 50.000 e 40.000 anni fa.
Le raccolte di manufatti del Castelperroniano (CP) trovati nella Francia centrale e nella Spagna settentrionale sono databili in questo intervallo di tempo. Fino ad ora, è il solo tipo di collezioni che ha consentito di associare i resti Neandertal con manufatti di stile UP. I manufatti CP includono anche ornamenti del corpo, praticamente sconosciuti in tutto il mondo Neandertal. Tuttavia, si è sostenuto che i manufatti CP non siano stati fabbricati da Neandertaliani, ma piuttosto i processi di formazione dei siti e la commistione degli strati abbiano portato all'associazione casuale di resti di Neandertaliani, assemblaggi CP, e gli ornamenti del corpo.
Fin dal Paleolitico medio, la pece di betulla fu largamente utilizzata dagli Uomini di Neanderthal come collante. Uno studio del 2019 ha dimostrato come queste prime produzioni di pece fossero realizzate attraverso una tecnica relativamente semplice, utilizzando dei focolari a cielo aperto.
Gli autori pubblicano una serie di datazioni al radiocarbonio ottenute con spettrometria di accelerazione di massa analizzando collagene osseo ultrafiltrato estratto da 40 frammenti di ossa ben conservate del tardo Musteriano, CP, e dagli strati Protoaurignaziani nel sito di Grotte du Renne (a Arcy-sur-Cure, Francia). I risultati sono incompatibili con l'ipotesi di commistione. Inoltre, si riporta una data direttamente calcolata su di un frammento di osso compatto di tibia di Neanderthal rinvenuto a Saint-Césaire (Francia). Questa data conferma l'assegnazione di manufatti CP agli ultimi Neanderthal dell'Europa occidentale. Inoltre, ed è ancora più importante, i dati stabiliscono che la produzione di ornamenti corporei nel CP è successiva all'arrivo degli umani moderni nelle regioni adiacenti dell'Europa. Questo nuovo comportamento potrebbe quindi essere il risultato di una diffusione culturale dai gruppi moderni ai gruppi di Neandertaliani.
Distribuzione territoriale
L'uomo di Neandertal è un genere originario dell'Europa occidentale e centrale, poi emigrato sulla via del Medio Oriente, sugli attuali territori di Iraq, Siria e d'Israele, con pochi individui fino in Asia centrale (Uzbekistan) e in Siberia. L'evoluzione che ha condotto alla comparsa dell'Homo neanderthalensis, o «neandertalizzazione», è stata lenta e progressiva, da gruppi europei isolati (Homo erectus, Homo georgicus, Homo antecessor). Può essere seguita partendo dai pre-Neandertaliani e a seguire fino ai Neandertaliani recenti.
- I pre-Neandertaliani antichi
La prima tappa corrisponde a fossili generalmente attribuiti a Homo heidelbergensis, possibile antenato, secondo alcune teorie anche dei sapiens moderni: è il caso dell'Uomo di Tautavel (-400.000 anni), rinvenuto in Francia, della mandibola di Mauer (-600.000 anni), trovato vicino a Heidelberg in Germania, o del cranio di Petralona (Grecia).
- I pre-Neandertaliani recenti
La prima tappa corrisponde ai fossili di Swanscombe (Inghilterra), di Steinheim (Germania) o della Sima de los Huesos à Atapuerca (Spagna).
- I Neandertaliani antichi
I successivi fossili con tratti innegabilmente Neandertaliani hanno un'età compresa tra -250.000 e -110.000 anni. Si può citare il cranio Biache-Saint-Vaast (Passo di Calais), i resti de la Chaise (Charente), la mandibole di Montmaurin (Alta-Garonna), i crani italiani di Saccopastore (Lazio) o l'abbondante materiale di Krapina in Croazia.
- I Neandertaliani classici
I Neandertaliani tipici, con caratteri derivativi più marcati hanno un'età compresa tra -100 000 anni e -28 000 anni, data degli ultimi rinvenimenti fossili, e presumibilmente della loro sparizione.
Inoltre, oltre ai fossili di Neandertal stesso (circa -42.000 anni), si ricordano gli scheletri di La Chapelle-aux-Saints, di Moustier, di La Ferrassie, di La Quina, di Saint-Césaire nel sud-ovest della Francia o della Spy in Belgio.
Gli ultimi Neandertaliani noti sono stati rinvenuti in Portogallo, in Spagna (Zafarraya -30.000 anni), in Croazia (Vindija, -32.000 anni) e nel nord-ovest del Caucaso (Mezmaiskaya, -29.000 anni). Come già visto queste date sono da considerare con precauzione, e sono spesso controverse.
Alcune ricerche condotte dal 1999 al 2005 sulla grotta di Gorham a Gibilterra suggeriscono che i Neandertaliani l'abbiano abitata dai -28.000 ai -24.000 anni. Dunque una lunga coabitazione con Homo sapiens, geograficamente presente dai -32.000 anni. Joao Zilhao, dell'università di Bristol critica fermamente questi ultimi risultati spostando all'indietro la data di estinzione a 37.000 anni fa. Le diverse teorie sulle date di estinzione e coabitazione con H.s.s. allo stato attuale non possono che definirsi in fase di sviluppo e verifica, ciò anche alla luce di sempre nuovi ritrovamenti, anche di terzi coabitatori cogeneri, nella fascia temporale dei 30.000 - 50.000 anni fa.
Antropologia fisica
Comparati ai moderni esseri umani i Neandertal erano di costituzione più robusta e avevano caratteri morfologici distintivi, specie per quanto riguarda il cranio, che gradualmente accumulava più aspetti derivati (in senso cladistico), specialmente in regioni geografiche relativamente isolate. Anche qui è da notare che le popolazioni anatomicamente moderne del paleolitico medio (vicino oriente come a Skul e Qafzeh), erano ugualmente massicce. Si pensa che la loro statura relativamente alta ma robusta sia un adattamento al clima freddo dell'Europa durante il Pleistocene, (legge di Allen) anche se in realtà questi tratti paiono già presenti in periodi caldi.
Craniometria
Un'accurata statistica, basata sull'analisi multivariata di forma e dimensioni del cranio, tuttavia, pur rilevando un habitus tipico del Neandertal, mostra sorprendenti vicinanze con crani attuali di forme estreme (C.Stringer, del British Museum).
Pigmentazione
Da uno studio del 2001 alcuni commentatori speculano sul fatto che i Neandertal dimostrino un tipo a capelli rossi, condividendone l'eredità con uomini attuali di tipo lentigginoso e di pelo rosso; in ogni caso altri ricercatori dell'epoca dissentivano. Gli studi molecolari del 2007 citati nel paragrafo iniziale Sintesi dell'aspetto esteriore sciolgono comunque ogni dubbio, ovvero sulla base dello studio sulle varianti dei melanocortin 1 receptor (mc1r), (o melanocyte-stimulating hormone receptor (MSHR), melanin-activating peptide receptor, o melanotropin receptor) una classe di recettori cellulari, recettori accoppiati a proteine G, che sono ligandi di ormoni peptidici Ipofisari, si deduce che la ridotta funzionalità di questi porti a sviluppare un fenotipo poco pigmentato a pelle chiara e capelli rossi. Lo studio è stato condotto, con tecniche di amplificazione genica su un gene responsabile della pigmentazione umana, appunto quello relativo al melanocortin 1 receptor (mc1r), che codifica sette proteine eterometriche transmembrana GTP-ligande (G protein)coupled receptor (GPCR). I capelli rossi e la pelle chiara risultano da alleli che comportino complete o parziali perdite di funzionalità dell'MC1R umano (huMC1R), alterando il bilancio nella sintesi eumelanina-feomelanina. I campioni provenivano da DNA ben conservato (sulla base di dati relativi ad abbondante contenuto amminoacidico e alla sua scarsa racemizzazione) di un esemplare dei Monti Lessini (in Italia) e dell'esemplare El Sidrón 1252 (Cueva de El Sidrón, in Spagna nelle Asturie).
Sviluppo fisico
Un'altra differenza secondo i paleoantropologi, potrebbe essere stata nella crescita, infatti si ritiene, analizzando la crescita del primo molare comparata con quello della dentatura di Homo sapiens, che i Neandertal crescessero più velocemente di quanto facciano gli uomini moderni, raggiungendo l'età adulta già a 15 anni. Questa crescita rapida, stando a quanto riportano gli esperti, è una caratteristica tipica dei primi ominidi. Tempi di sviluppo più rapidi, secondo gli studiosi, sembrano essere stati una necessità per una specie che viveva in terre molto fredde e inospitali, dove la mortalità infantile era molto elevata e l'aspettativa di vita breve.
Ibridazione con Homo sapiens
Nel corso della seconda metà del XX secolo si è assai dibattuto in ambito accademico se l'uomo di Neandertal sia da considerare una sottospecie estinta di Homo sapiens (denominata Homo sapiens neanderthalensis), con la conseguente possibilità di incrocio e ibridazione tra i due, oppure se sia una specie autonoma (Homo neanderthalensis). Comunque sia, i due gruppi convissero per migliaia di anni in Europa e Asia Minore. Indagini genetiche eseguite nel 2008 su DNA fossile recuperato su alcuni resti di uomo di Neandertal, sembravano indicare che i due tipi umani parzialmente contemporanei appartenessero a due diverse specie, tuttavia più recenti scoperte riguardanti il genoma umano sembrerebbero in grado di accertare che neanderthalensis e sapiens apparterrebbero alla stessa specie e che le differenze anatomiche e somatiche sarebbero dovute a cause climatiche, ambientali, morfologiche e di alimentazione.
Analisi del DNA
Il biologo molecolare svedese Svante Pääbo ha testato almeno 70 campioni provenienti da Neandertaliani prima di trovarne uno sufficiente per le analisi del DNA; inizialmente sequenze da ossa di 38.000 anni provenienti dal sito croato di Vindija mostrano una condivisione del 99,5% del DNA con sapiens: i due appartenenti al genere Homo da ciò divergerebbero da circa 500.000 anni. Un articolo sulla rivista Nature stima la divergenza a 516.000 anni, dove le prove paleontologiche, sufficientemente concordi, mostrano un periodo di 400.000 anni. Svante Pääbo ha proposto un progetto per la ricostruzione totale del patrimonio genetico di Homo neanderthalensis. Edward Rubin del Lawrence Berkeley National Laboratory di Berkeley, California dimostra che il genoma dei Neandertal è simile al nostro dal 99,5 al 99,9%.
Nel novembre 2006, un nuovo articolo viene pubblicato sulla rivista scientifica Proceedings of the National Academy of Sciences, dove un gruppo di ricerca europeo ripropone il tema dell'ibridazione. Contemporaneamente il 16 novembre 2006 Science Daily pubblica la tesi opposta, elaborata dal Berkeley Lab e dal Joint Genome Institute, secondo cui non vi sarebbero evidenze di incroci tra i due gruppi umani e la sovrapposizione genetica non supererebbe il 99,5%.
Edward Rubin, direttore di entrambi gli istituti, conclude:
«Anche se non siamo in grado di affermare definitivamente che l'ibridazione tra le due specie non abbia avuto luogo, l'analisi del DNA nucleare del Neandertal suggerisce una bassa probabilità che questo sia avvenuto a livello apprezzabile.» La più accurata analisi molecolare condivisa a livello internazionale e attualmente confermata, stima che l'Homo sapiens e l'Homo neanderthalensis si separarono non più di 800.000 anni fa e non vi sono ancora prove evidenti accettate dalla maggioranza della comunità scientifica su un eventuale ricongiungimento. Attualmente la comunità scientifica, sulla base di alcuni studi di Svante Pääbo dell'Istituto Max Planck di Lipsia, considera comunque possibile tra le varie ipotesi, un ricongiungimento genetico tra le due specie, sulla base della presenza, nel genoma di parte della popolazione umana attuale (esclusi gli africani), di circa l'1-4% di materiale genetico compatibile con quello neandertaliano.
In un ulteriore articolo di Svante Pääbo, in collaborazione con il Dipartimento di Genetica di Harvard e il MIT, viene proposta una datazione dell'ultimo periodo di scambio di geni tra Neandertal e umani moderni (Homo sapiens sapiens).
Una delle scoperte chiave dell'analisi del genoma di Neandertal è che i Neandertal condividono più varianti genetiche con non-africani che con gli africani. Questa osservazione è in linea con due ipotesi:
1) l'incrocio tra Neandertal e gli esseri umani moderni è avvenuto dopo che gli umani moderni sono usciti dell'Africa o
2) la struttura delle popolazioni degli antenati di Neandertal e di quelli degli esseri umani moderni erano in parte simili.
Queste ipotesi possono fare previsioni diverse sulla data dell'ultimo scambio di geni tra gli antenati di Neandertal e gli umani moderni non africani. Gli autori stimano questa data misurando il grado di linkage disequilibrium (LD) (spesso definita "associazione allelica"; particolari alleli in due o più loci vicini mostrano associazione allelica se si verificano con frequenze significativamente diverse da quelle previste in base alle frequenze alleliche dei singoli) tra il genoma degli attuali europei e trovano che l'ultimo flusso genico da Neandertal agli europei si è probabilmente verificato 37.000-86.000 anni fa (BP), e più probabilmente 47.000-65.000 anni aC. Ciò supporta l'ipotesi di un incrocio recente e suggerisce che l'incrocio si è verificato quando gli esseri umani moderni portatori delle tecnologie del Paleolitico superiore incontrarono i Neandertal a seguito della loro espansione fuori dall'Africa.
Le tracce neandertaliane più recenti
Si dibatte lungamente sugli ultimi rappresentanti noti dei nostri parenti più prossimi. Quest'argomento, assieme al mistero della scomparsa della popolazione neanderthaliana, è forse uno dei più controversi della scienza paleoantropologica e il più soggetto a evoluzione. Le datazioni tramite spettrometria di massa da parte di Fred Smith ed Erik Trinkaus, (Northern Illinois University e Washington University rispettivamente), da scavi di Vindija (Croazia) portano a 28.000 anni i reperti più recenti, con articoli pubblicati a cavallo del 2000. Nuove datazioni ricollocano indietro a 32.000 anni i reperti. Vengono scoperti altri fossili recenti sulle coste atlantiche del Portogallo, e si riscontrano evoluzioni morfologiche verso una maggiore modernità. Contemporaneamente viene messa in discussione l'associazione di alcune culture litiche (Aurignaziano, Musteriano) ai vari Homo.
La scomparsa dell'intera popolazione neanderthaliana
La teoria del "rimpiazzamento"
Si dibatte sulla completa scomparsa della popolazione, non perfettamente spiegabile sulla base delle sole caratteristiche fisiche degli individui. Si trattava di una specie lungamente adattata all'ambiente colonizzato, con un volume cranico pari o superiore ai sapiens attuali, e di cultura tecnica almeno inizialmente sovrapponibile nelle due popolazioni. Le prime ipotesi teorizzate parlano di lenta ibridazione con Sapiens moderni, eliminazione fisica (genocidio), competizione, o selezione sessuale. La difficoltà ad analizzare lo scarso materiale genetico sopravvissuto completa il quadro. Certamente la lunga coesistenza di uomo di Neandertal e uomo moderno pongono sul tavolo della discussione molti problemi irrisolti e l'argomento è in costante riscrittura. Nel 2005 sul Journal of Economic Behaviour and Organization Jason Shogren, economista dell'Università del Wyoming di Laramie, pubblica con i suoi collaboratori un articolo in cui avanza una teoria sulla scomparsa dell'uomo di Neandertal. Lo studioso avanza l'ipotesi che Homo neanderthalensis si sia dovuto scontrare con la particolare cultura dell'Homo sapiens: questa cultura si basava su tecniche avanzate di commercio, cosa che portava più tempo libero rispetto a una cultura basata sulla caccia. Il tempo libero ottenuto avrebbe permesso lo sviluppo di specializzazioni non strettamente legate alla sussistenza, come costruire utensili sempre più complessi o dedicarsi all'arte. La complessità e la versatilità di una tale cultura avrebbe avuto esito fatale per la più "tradizionale" cultura dei Neandertal. Stephen Kuhn e Mary Stiner dell'università dell'Arizona, sulla rivista Current Anthropology, propongono, documentandola, la tesi per cui la principale causa di estinzione fu la mancata suddivisione dei lavori tra i sessi. I più organizzati sapiens, più efficientemente, poterono competere affidando alle donne compiti stanziali, e meno gravosi, affidando ai maschi i ruoli di cacciatori e approvvigionatori di materiali. La prole, protetta e anch'essa stanziale, avrebbe avuto più possibilità di sopravvivenza. Un'altra ipotesi sulla loro estinzione, fa riferimento invece ai drastici cambiamenti nella composizione della magnetosfera terrestre, dovuti all'azione di forti variazioni del campo magnetico terrestre, con conseguente variazione della radiazione solare che avrebbe comportato, a sua volta, mutazioni negative per la specie, fino a portarla all'estinzione. Due caratteristiche dei neanderthalensis vanno rimarcate. La prima è che il loro fisico era strutturato per esprimere al meglio la forza, mentre quello di alcuni, ma non tutti, i sapiens loro contemporanei come i Cromagnonoidi, a gambe più lunghe e a bacino più stretto e compatto, privilegiava le capacità di resistenza nella corsa. Quindi quei sapiens sarebbero stati più resistenti nel percorso di sensibili distanze con quel tipo di andatura, ma svantaggiati nella forza fisica e nella lotta. Una caratteristica che si pensava differenziasse sapiens e Neandertal, la diversa alimentazione, con sapiens marcatamente onnivori e Neandertal carnivori si è variamente rivelata parziale e dipendente esclusivamente da singole situazioni. Entrambi ad esempio erano specie ben adattate agli ambienti costieri con un'alimentazione basata su frutta e verdura, prodotti della pesca, raccolta di molluschi e caccia. Soggetta a periodiche discussioni è anche la capacità dei Neandertal di cacciare grosse prede (come la megafauna); mentre esistono prove che anche i più antichi esponenti della nostra specie utilizzassero zagaglie, e altri ordigni da lancio, non si sa se i Neandertal utilizzassero armi da lancio. Avrebbero cioè dovuto approcciarsi alle prede (e a eventuali nemici) a distanza ravvicinata. I Neandertal, almeno alcune popolazioni, erano comunque prettamente carnivor e superpredatori.
La teoria dell'"ibridazione"
Uno studio pubblicato su Science Advance nel 2020 afferma che circa 50.000 anni fa, mentre Homo sapiens si diffondeva fuori dell'Africa, avvennero accoppiamenti fertili con popolazioni neandertaliane e denisoviane sino al loro assorbimento e sostituzione con popolazioni tipicamente sapiens ma nel cui genoma rimase traccia dell'avvenuta mescolanza genetica. Nel 2010 sono stati pubblicati i risultati di una ricerca quadriennale che mirava a mappare il genoma dei Neanderthal, è stato comparato del DNA raccolto da dei fossili di Neanderthal con del Dna degli umani contemporanei, e i risultati hanno stupefatto la comunità scientifica, nel DNA unico condiviso delle moderne popolazioni una porzione rilevante tra 1 e il 4% era costituito da DNA Neanderthaliano.
Ricostruzioni dell'aspetto, secondo la scienza e gli stereotipi
Molto ma non tutto si può dire del vero aspetto di questo ominide ricostruendolo dai suoi resti fossili. La scienza contemporanea ha messo a punto varie tecniche per la ricostruzione delle parti non fossilizzabili, i tessuti molli, a partire da elementi quali le inserzioni tendinee sulle ossa, le linee di forza sulle stesse, i livelli di consunzione dei denti e altro ancora. La biologia molecolare ha poi fornito ulteriori elementi per valutare l'espressione genica di caratteri non conservabili, appunto come il colore dei capelli. Il Neandertal possedeva la tecnologia necessaria a confezionare indumenti; a tale scopo utilizzava prevalentemente pelli, per la concia delle quali costruiva utensili quali i raschiatoi musteriani. Le zone climatiche frequentate imponevano sicuramente l'uso di coperture, e possiamo rappresentare il Neandertal tipico prevalentemente vestito.
Oltre a ciò si segnala la capacità simbolica e artistica, che ha portato all'uso, almeno episodico, di monili e pendagli. In passato, al contrario, è spesso stato rappresentato come lo stereotipo dell'uomo cavernicolo, tale ricostruzione ripete né più né meno la figura dell'uomo selvatico presente nella tradizione popolare europea e raffigurata sin dal Medioevo. Secondo alcune interpretazioni di studiosi di criptozoologia, non di antropologi o paleontologi, poteva però essere di aspetto scimmiesco, dotato di un fitto villo pilifero distribuito su tutta la superficie del corpo che lo proteggeva dal clima freddo europeo.
Un'immagine da alcuni supposta e interpretata di Neandertal è una rappresentazione preistorica nella caverna di Isturits sui bassi Pirenei francesi, frequentata da 80.000 a 10.000 anni fa prima dai Neandertal e poi dai Cro-magnon. Vi compare la rappresentazione di una testa-trofeo di un essere antropomorfo con testa ovale senza fronte e mento, collo grosso, occhi a mandorla infossati sotto grandi arcate frontali, con un grosso naso, orecchie piccole, con capelli corti a spazzola e dritti a cresta sulla testa che proseguivano sulla schiena in una sorta di criniera equina, e peli fitti simili su quasi tutta la faccia e collo simili al manto di un cervo.
Storia dei principali reperti fossili europei
I resti rinvenuti nel 1856 da Johann Fuhlrott nella valle di Neander consistevano nella parte superiore del cranio, alcune ossa, parte dell'osso pelvico, alcune costole, e ossa del braccio e della spalla. In precedenza erano stati scoperti altri fossili, infatti già nel 1829 nel Belgio venne trovato parte di un cranio di un bambino di due anni e mezzo. Questi, però, venne riconosciuto come arcaico soltanto nel 1836. Nel 1848 a Gibilterra venne trovato un cranio adulto, poi battezzato Gibraltar 1, ma la sua esistenza rimase sconosciuta alla scienza fino al 1864, quando venne riconosciuto come appartenente a un individuo Neandertal Altri due scheletri di Homo neanderthalensis, di almeno 60.000 anni, vennero trovati in Belgio nel 1886 da Marcel de Puydt e Max Lohest. Altri rinvenimenti importanti vennero fatti in Croazia nel 1899 da Dragutin Gorjanovic-Kramberger e nel 1908 in Francia a La Chapelle-aux-Saints da Jean Bouyssonie che rinvenne lo scheletro di un uomo anziano, risalente a 50000 anni, il cui volume cranico era di ben 1620 centimetri cubi.
Nel 1939 venne rinvenuto nella grotta Guattari a San Felice Circeo, un cranio presumibilmente appartenente a Homo neanderthalensis.
Nel secondo dopo guerra emersero ancora altri resti importanti; tra il 1953 e il 1960 nella grotta di Shanidar in Iraq vennero scoperti 9 scheletri di uomini di Neandertal, risalenti a un periodo compreso tra i 70 e i 40.000 anni fa, e nel 1979 nel villaggio di Saint-Césaire in Francia uno scheletro completo risalente a 35.000 anni fa.
Nel 1868 a Cro-Magnon in Francia vennero trovati da alcuni operai i resti di un uomo risalenti a 28.000 anni fa; era venuto alla luce uno dei più antichi progenitori della nostra specie (Homo sapiens). Era un rappresentante della nostra specie umana che, proveniente dall'Africa o dall'Asia, migrando stava insediandosi in Europa, confinando verso la penisola iberica gli ultimi Neandertal.
Uomo di Cro-Magnon
L'uomo di Cro-Magnon è un'antica forma di Homo sapiens, ascrivibile a popolazioni umane moderne, largamente diffusa nel paleolitico superiore in Europa. È rappresentato da quattro scheletri provenienti dal riparo sottoroccia di Cro-Magnon, rinvenuti nel 1868 presso Les Eyzies-de-Tayac-Sireuil in Dordogna e da sette scheletri raccolti nelle Grotte dei Balzi Rossi (Liguria), definiti a suo tempo come cromagnonoidi.
Antropologia fisica
Antropologicamente si osserva una certa stabilità delle caratteristiche cromagnonoidi che sono essenzialmente di tipo caucasoide:
- alta statura (media 1,75 m per gli uomini, con punte oltre 1,95 m) con gambe lunghe e braccia corte;
- faccia larga e bassa con cranio lungo dalla fronte all'occipite (dolicocefalia e cameprosopia), spesso denotata come disarmonica;
- orbite basse e rettangolari;
- naso prominente e spesso aquilino;
- grande capacità cranica (1.650 cm³).
Genetica
Dalle moderne indagini genetiche sembra potersi affermare che i cromagnonoidi entrarono in Europa dall'Asia centrale verso il 30.000 a.C., portando il particolare marcatore genetico M173, derivato da M45, che pare fosse diffuso in popolazioni asiatiche del paleolitico da cui sarebbero derivate anche alcune popolazioni siberiane e amerinde (marcatore M242 e discendenti). Indagini genetiche effettuate su scheletri paleolitici di Cro-Magnon sparsi in Eurasia, rivelano che l'aplogruppo mitocondriale predominante fosse l'U, specialmente nelle sue cladi U2, U5 e U8.
Nella grotta di Fumane in Veneto, un individuo appartenente alla cultura aurignaziana è risultato positivo all'aplogruppo mtDNA R*, così come l'uomo rinvenuto a Ust'-Iim nella Siberia occidentale. Uno scheletro appartenente alla cultura maddaleniana e rinvenuto nella grotta di La Pasiega, in Cantabria, risulta portare l'Aplogruppo mtDNA H. L'aplogruppo mitocondriale estratto dalla mandibola di Oase I, rinvenuta a Pe?tera cu Oase in Romania, è stato identificato come N.
La sequenza del DNA mitocondriale eseguita nel 2003 da un team italo-spagnolo diretto da David Caramelli, su due fossili di Cro-Magnon identificati come Pelosi 1 e 2, risalenti rispettivamente a 23 e 24.000 anni fa, ha identificato il DNA mitocondriale come aplogruppo N. Questo aplogruppo si ritrova nelle moderne popolazioni del Medio Oriente, Nord Africa e Asia centrale, mentre gli aplogruppi derivati si trovano anche in popolazioni eurasiatiche, polinesiane e tra i nativi americani.
Secondo Bryan Sykes professore di genetica umana presso l'università di Oxford, nel suo libro pubblicato nel 2001 "The Seven Daughters of Eve", l'uomo di Cro-Magnon si sarebbe accoppiato con sette tipi di donne successivamente, che in base al loro DNA mitocondriale vengono nominate: Ursula (aplogruppo U) trovata in Siberia, Xenia (aplogruppo X), Tara (T) e Helena (H) trovate in Europa nel paleolitico, e poi Katrine (K) e Velda(V) evolutesi nel mesolito e infine Jasmine (J) venuta dal levante nel neolitico, per dare origine ai principali aplogruppi mitocondriali diffusi nelle popolazioni europee moderne.
Per quanto riguarda gli aplogruppi del cromosoma Y, si hanno a disposizione meno dati visto che solo gli individui di sesso maschile ne possiedono uno e inoltre si degrada più facilmente nel tempo rispetto all'mtDNA. L'Uomo di Ust'-Iim in Siberia portava l'aplogruppo Y-DNA K, risultando negativo a tutti i polimorfismi a singolo nucleotide che contraddistinguono i suoi aplogruppi derivati, da L a T, oggi maggiormente diffusi. Sempre in Siberia, è stato rinvenuto uno scheletro di un bambino di 3-4 anni appartenente alla Cultura di Mal'ta-Buret', il cui aplogruppo Y-DNA è risultato essere l'R*, antenato dei moderni R1 (da cui R1a e R1b, gli aplogruppi più diffusi in Europa) e R2. Lo scheletro rinvenuto a Kostenki, nella Russia europea, portava l'aplogruppo Y-DNA C1. La mandibola di Oase 1 ritrovata in Romania risultò invece appartenere all'aplogruppo F, antenato degli aplogruppi Y-DNA che rappresentano il 90% della popolazione mondiale.
Analisi genetiche effettuate sugli scheletri dei cromagnoidi cacciatori-raccoglitori mesolitici della cava di Loschbour in Lussemburgo e Motala in Svezia hanno rivelato che gli uomini di Cro-Magnon hanno contribuito anche al genoma autosomico degli odierni Europei, assieme agli agricoltori neolitici e un terzo ceppo ancestrale correlato ai Siberiani paleolitici. Attualmente, le popolazioni europee che hanno più DNA autosomico correlato ai cacciatori-raccoglitori europei (Cro-Magnon) sono gli abitanti dei Paesi Baltici.
Vita dei Cro-Magnon
I Cro-Magnon avevano una dieta di carne, grano, carote, cipolle, rape ed altri alimenti; nel complesso, una dieta molto bilanciata.
Tra gli artefatti Cro-Magnon giunti fino a noi vi sono capanne, pitture murali, incisioni.. Le capanne erano costruite in roccia, argilla, ossa, rami e pelo di animali.
I Cro-Magnon utilizzavano manganese e ossido di ferro per le loro pitture rupestri, e potrebbero aver creato, circa 15.000 anni fa, il primo calendario.
I Cro-Magnon devono essere entrati in contatto con gli uomini di Neanderthal e sono spesso indicati come la causa dell'estinzione di questi ultimi; in realtà, sembra che umani moderni dal punto di vista morfologico abbiano convissuto con i Neanderthal per circa 60.000 anni nel Levante, e per più di 10.000 anni in Francia.
Culture preistoriche associate ai cro-magnon
Prima della comparsa dell'uomo moderno in Europa esisteva la cultura Musteriana, la caratteristica industria litica prodotta, sia dagli uomini di neanderthal, sia dai primi uomini moderni, almeno fino a 30.000 anni fa. Da 40.000 anni fa si sovrappone il Castelperroniano con un'industria litica più diversificata ed evoluta, che potrebbe interpretarsi con la prima comparsa in Europa di una razza umana moderna. L'Aurignaziano è la prima cultura preistorica in cui compare l'arte figurativa, ed è chiaramente originaria in Siria per poi estendersi progressivamente prima ai balcani poi nell'occidente del Mediterraneo e infine penetrare nella Europa centrale, per cui chiaramente è stata portata da popolazioni distinte dai cro-magnon, infatti i reperti antropologici associati nelle cave in Moravia e Romania mostrano una razza umana moderna ma di struttura ancora arcaica. La fase successiva vede il progressivo raffreddamento del clima in Europa, mentre fanno ingresso attraverso la steppa ucraina provenienti dall'Asia centrale i cro-magnon. Con l'estendersi dei ghiacciai in Germania e sulle Alpi si ebbe la separazione tra la zona dell'Europa occidentale dall'Europa orientale. Mentre in Europa orientale l'industria litica si semplifica all'essenziale durante il Gravettiano tra 30.000 e 10.000 anni fa, e testimonia la vita nomade dei cacciatori cro-magnon da cui discenderanno le popolazioni indoeuropee moderne. In Europa occidentale, e principalmente in Francia e nella penisola Iberica, si sviluppano le massime industrie del Paleolitico superiore: inizia da 21.000 anni fa la cultura Solutreana a cui segue la Magdaleniana tra 18.000 fino a 10.000 anni fa, che ci ha lasciato le meravigliose pitture rupestri naturalistiche delle grotte.
Uomo di Oberkassel
Nella località di Oberkassel, presso Bonn in Germania, sono stati ritrovati nel 1914 due scheletri in una doppia sepoltura, datati al 10.000 a.C. - 15.000 a.C. e riferibili al Maddaleniano. Si tratta di uno scheletro maschile e di uno femminile assai diversi tra loro. Caratteristico appare specialmente il cranio maschile molto capace (1.600 cm³) leggermente dolicocefalo, con faccia fortemente cameprosopa e orbite molto basse, in qualche modo accentuando la disarmonia di Cro-Magnon.
Il cranio della donna è più alto e più stretto e non è evidentemente cromagnonoide, ma ricorda invece il tipo di Brünn. La statura è di 166 cm nell'uomo e 147 cm nella donna.
Diffusione geografica e varietà locali
- le popolazioni di Mechta-Afalou in Nord-Africa (Berberi),
- la popolazione maglemosiana (proto-nordici della varietà dalo-falica) in Scandinavia,
- le popolazioni neolitiche delle culture del Dneper-Donets e di Sredny-Stog (forse i proto-Indoeuropei) nella Russia meridionale,
- i Guanci delle isole Canarie, ormai estinti, probabilmente discendenti dei Berberi,
- i nativi americani Dakota in Nordamerica,
- l'uomo del Carmelo.
Non è chiaro come i Cro-Magnon abbiano contribuito alla genetica delle popolazioni odierne in Asia, ma è stato rilevato che in Asia i portatori delle culture siberiane Afanasevo e Tagar erano essenzialmente cromagnonoidi.
 datate approssimativamente 16.000 anni fa (Paleolitico superiore).jpg)
Pitture rupestri di cro-magnon delle grotte di Lascaux (Francia) datate approssimativamente 16.000 anni fa.
Homo sapiens
Homo sapiens (Linnaeus, 1758) è la definizione tassonomica dell'essere umano moderno. Appartiene al genere Homo, di cui è l'unica specie vivente, alla famiglia degli ominidi e all'ordine dei primati. L'epoca che va dal periodo interglaciale medio, circa 300.000 anni fa, all'epoca odierna, vede la comparsa in Africa orientale e la diversificazione della specie Homo sapiens. Secondo le teorie prevalenti, dal continente africano, circa 65-75.000 anni fa, parte della specie iniziò un percorso migratorio che, attraverso un corridoio mediorientale, la portò a colonizzare l'intero pianeta. La precisa datazione dei primi esemplari definibili sapiens, tradizionalmente posta a circa 130.000 anni fa, è stata spostata dalle scienze paleontologiche più indietro nel tempo, grazie a ritrovamenti nei tufi vulcanici della valle del fiume Omo in Etiopia. Per mezzo di tecniche basate sui rapporti fra gli isotopi dell'argon, alcuni reperti anatomicamente simili all'uomo moderno sono stati datati a 195.000 anni fa, con un'incertezza di ±5.000 anni. Nuove datazioni del 2017 su reperti rinvenuti nel 1961 nel sito archeologico di Jebel Irhoud, in Marocco, sposterebbero l'origine dell'Homo sapiens a circa 300.000 anni fa.
Le caratteristiche anatomiche che differenziano maggiormente Homo sapiens dalle specie a esso affini e le conseguenze sulla sua biologia e comportamento si concentrano principalmente sul grado di sviluppo del sistema nervoso, inteso come massa e complessità, e sulla sua organizzazione neurale.
Sistema neuro sensoriale e adattamento
Gli umani hanno un cervello molto strutturato e sviluppato, molto grande in proporzione alle dimensioni dell'individuo (quoziente di encefalizzazione), con notevoli doti di neuroplasticità, capace di un pensiero sviluppato sotto forma di creatività, ragionamento astratto, linguaggio e introspezione. Questa capacità mentale li distingue dagli altri esseri viventi. Altre caratteristiche peculiari sono:
- La postura eretta, rara nei mammiferi e presente solo nei canguri.
- Gli arti superiori liberi e con notevoli doti di precisione, sensibilità e prensili per l'origine arboricola (caratteristica comune a tutto l'ordine tassonomico).
- La visione binoculare necessaria alla percezione della profondità (stereopsi).
- La conseguente precisione nella manipolazione degli oggetti.
Tali caratteristiche hanno permesso agli esseri umani di creare una grande varietà di manufatti e utensili atti a migliorare il proprio adattamento all'ambiente, la sopravvivenza e l'espressione creativa.
Origine e diffusione
Le più recenti pubblicazioni accademiche, datate 10 luglio 2019 (Università di Tubinga pubblicati su Nature), riportano che il più antico esemplare in Eurasia ad oggi noto di Homo sapiens è stato scoperto in una grotta, denominata Apidima, del Peloponneso (Grecia) nelle vicinanze di Kalamata e Sparta. I ricercatori scoprirono circa 20.000 reperti (ossa, denti, strumenti litici e altro, tra cui due crani, denominati Apidima 1 e Apidima 2); il cranio "Apidima 1" è la più antica testimonianza a oggi ritrovata di Homo sapiens, datato ad oltre circa 210.000 anni fa. Apidima 1 è stato datato col metodo scientifico attualmente più evoluto basato sull'Uranio-Torio-230 ed è risultato più antico di decine di migliaia di anni rispetto agli antecedenti reperti di Homo sapiens con datazione certa ritrovati in Africa o Europa, precedentemente considerati i più vecchi, con un salto cronologico impressionante che ridiscute tutta la teoria del Sapiens. La ricercatrice a capo del progetto Katerina Harvati spiega che almeno due popolazioni vivevano in Grecia meridionale nel Pleistocene medio: una primigenia popolazione di Homo sapiens, seguita dalle evidenze dei ritrovamenti dopo circa 40.000 anni da una popolazione di neandertaliani. Al 2021 sono in corso le estrazioni del DNA dagli antichissimi ritrovamenti, anche se, secondo lo stesso Team, l'operazione potrebbe essere difficilissima.
Dall'origine ipotizzata di provenienza africana della specie ("Out of Africa Hypothesis", ipotesi nella quale l'uomo sarebbe nato in Africa e successivamente in espansione nel resto del mondo), nei ritrovamenti etiopici in tufi vulcanici della valle del fiume Omo, si possono datare con tecniche basate sui rapporti isotopici dell'argon, a 195.000 anni (con un'incertezza di ±5 000 anni) due esemplari (Omo II e Omo I), rispettivamente di Homo erectus moderno e Homo sapiens arcaico (da cui secondo quest'ipotesi si sarebbero evoluti gli uomini attuali); ad oggi, Homo sapiens si è diffuso su tutta la superficie delle terre emerse con una popolazione totale che ha superato, nel marzo 2017, i 7,4 miliardi di individui. Secondo ritrovamenti avvenuti in Marocco nel 2017, l'Homo sapiens avrebbe cominciato a diffondersi secondo lo studio di Daniel Richter e Shannon McPherron del Max Planck Institute, nell'intero continente africano già 315.000 anni fa, stimando la datazione del sito dei ritrovamenti.
Organizzazione sociale
Similmente alla maggior parte dei primati, Homo sapiens è un animale sociale. È inoltre particolarmente abile nell'utilizzo di sistemi di comunicazione per l'espressione, lo scambio di idee e l'organizzazione; crea complesse strutture sociali composte da gruppi in cooperazione e competizione, che variano dalle piccole famiglie e associazioni fino alle grandi unioni politiche, scientifiche, economiche. L'interazione sociale ha introdotto una larghissima varietà di tradizioni, rituali, regole comportamentali e morali, norme sociali e leggi che formano la base della società umana. La specie umana manifesta il desiderio di capire e influenzare il mondo circostante, cercando di comprendere, spiegare e manipolare i fenomeni naturali attraverso la scienza, la filosofia, la mitologia e la religione. Questa curiosità naturale ha portato allo sviluppo di strumenti tecnologici e abilità avanzate; Homo sapiens è l'unica specie ancora vivente che comunica attraverso la scrittura simbolica, utilizza il fuoco, cuoce i propri cibi, si veste ed usa numerose tecnologie. Gli esseri umani possiedono anche un marcato apprezzamento per la bellezza e l'estetica che, combinato col desiderio di auto-espressione, ha condotto a creative innovazioni culturali quali le arti, comprensive di tutte le discipline musicali, figurative e letterarie.
Evoluzione e diffusione della specie. La teoria attualmente più riconosciuta stima che:
- la sottotribù Hominina si sia evoluta nel Rift africano da una popolazione di Ominidi, progenitori comuni agli scimpanzé, circa 5-6 milioni di anni fa;
- il genere Homo si sia differenziato 2,3-2,4 milioni di anni fa dall'Australopithecus e diffuso sul globo come Homo erectus (Out of Africa I)
- la specie Homo sapiens si sia sviluppata anch'essa in Africa circa 300.000 anni fa e successivamente (100.000-65.000 anni fa) sia ugualmente migrata tra i continenti (Out of Africa II) con possibili ibridazioni successive con specie affini (Homo neanderthalensis, Homo di Denisova)
Evoluzione biologica
Lo studio scientifico dell'evoluzione umana comprende lo sviluppo del genere Homo e lo studio degli altri ominidi, quali ad esempio Australopithecus, a esso strettamente correlati. Gli umani moderni appartengono alla specie Homo sapiens, per alcuni autori suddivisibile in due sottospecie: Homo sapiens sapiens e Homo sapiens idaltu (tradotto approssimativamente come "uomo saggio maggiore"), estinto.
Dal punto di vista anatomico, gli umani moderni appaiono in testimonianze di reperti della grotta di Apidima in Grecia (Europa) risalenti a 210.000 anni fa e successivamente 80.000 anni più tardi in altri reperti risalenti 130.000 anni fa in Africa. Inoltre sono stati rinvenuti in tufi vulcanici della valle del fiume Omo in Etiopia resti risalenti a 195.000 anni fa, datati con tecniche basate sui rapporti isotopici dell'argon, con un'incertezza di ±5 000 anni.
I parenti più stretti ancora viventi di Homo sapiens sono le due specie appartenenti al genere Pan, comunemente noti come scimpanzé: il bonobo (Pan paniscus) e lo scimpanzé comune (Pan troglodytes). Le due specie sono ugualmente vicine, ovvero condividono lo stesso antenato comune; la differenza principale tra essi è l'organizzazione sociale: matriarcale per il bonobo e patriarcale per lo scimpanzé comune. Il sequenziamento completo del genoma ha portato alla conclusione che "dopo 6,5 milioni di anni di evoluzione separata, le differenze tra bonobo/scimpanzé ed umani sono soltanto dieci volte maggiori di quelle esistenti tra due persone qualsiasi e dieci volte minori di quelle esistenti tra ratti e topi".
Infatti, il 98,6% della sequenza di DNA è identica tra le due specie di scimpanzé e gli esseri umani. È stato stimato che la linea umana si sia distaccata da quella degli scimpanzé circa cinque milioni di anni fa e da quella dei gorilla circa otto milioni di anni fa. Tuttavia, un cranio ominide rinvenuto in Ciad nel 2001, classificato come Sahelanthropus tchadensis, risale approssimativamente a sette milioni di anni fa, la qual cosa potrebbe indicare una divergenza precedente; anche studi del 2009 su Ardipithecus ramidus portano a 5,4-7,4 milioni di anni la probabile divergenza. Queste minime differenze genetiche hanno portato alcuni scienziati, il più noto dei quali al vasto pubblico è Jared Diamond, a ipotizzare una riunificazione di uomini e scimpanzé sotto lo stesso genere Homo, come nell'originale schema di Linneo del Systema Naturae, edizione 1758. Ciò implicherebbe ovviamente una revisione totale almeno dei generi Pan, Ardipithecus, Kenyanthropus, Australopithecus e Homo.
L'attuale variabilità genetica della specie umana è estremamente bassa, comparativamente a quanto succede in altri raggruppamenti tassonomici animali.
I genetisti Lynn Jorde e Henry Harpending dell'università dello Utah hanno suggerito che la variazione del DNA umano è piccolissima se comparata con quella di altre specie e che durante il Tardo Pleistocene, la popolazione umana fosse ridotta a un piccolo numero di coppie genitoriali - non superiori alle 10.000 e forse intorno alle 1.000 - con la conseguenza di un pool genico residuo molto ristretto. Sono state formulate varie spiegazioni per questo ipotetico collo di bottiglia, tra cui la più famosa Teoria della catastrofe di Toba. L'evoluzione umana è caratterizzata da un certo numero di importanti tendenze fisiologiche, incluse l'espansione della cavità cerebrale e del cervello stesso, che arriva, con una distribuzione variabile per ogni singolo individuo, a un volume tipico di 1260 cm³, oltre il doppio di quello di uno scimpanzé o gorilla. Il ritmo di crescita postnatale del cervello umano differisce da quello delle altre scimmie antropomorfe (eterocronia), permettendo un lungo periodo di apprendimento sociale e l'acquisizione del linguaggio nei giovani umani. Gli antropologi sostengono che la riorganizzazione della struttura del cervello sia più importante della stessa espansione cerebrale. Altri significativi cambiamenti evolutivi includono una riduzione dei denti canini, lo sviluppo della locomozione bipede e la discesa della laringe e dell'osso ioide che permise il linguaggio. Come siano collegate queste tendenze e quale sia il loro ruolo nell'evoluzione di una complessa organizzazione sociale e della cultura rimangono questioni ancora dibattute.
Tipologia
L'Homo sapiens è una specie monotipica. Nel tempo sono state proposte specie, sottospecie e paleosottospecie di Homo sapiens, anche se sono in corso tutt'oggi dibattiti sull'argomento, dato che specie e sottospecie sono definizioni convenzionali e non differenziazioni oggettive calcolabili o rilevabili. Alcuni antropologi considerano la specie costituita da due diverse paleosottospecie: Sapiens e Idaltu. Molti considerano i Neandertal una specie ed altri una sottospecie. Ed infine alcuni considerano Homo heidelberg un progenitore, altri una specie ed altri ancora una paleosottospecie.
- Homo sapiens sapiens, l'essere umano moderno
- Homo sapiens idaltu, paleosottospecie estinta
- Homo sapiens heidelbergensis (uomo di Heidelberg)
- Homo sapiens neanderthalensis (uomo di Neanderthal)
Con l'utilizzo di tecniche di biologia molecolare per la verifica di eventuali riapparentamenti genetici, al 2011 gli studi basati sull'analisi matrilineare del DNA mitocondriale, mostravano una scarsa possibilità di passata ibridazione, mentre le analisi del genoma nucleare, anche stimolate dal progetto Neanderthal genome project del Max Planck Institute for Evolutionary Anthropology tedesco e del 454 Life Sciences statunitense di sequenziamento del genoma neandertaliano indicano una vasta ibridazione; la ricerca Archeogenetiche di Svante Pääbo (Max Planck Institute) ha dimostrato che circa il 70-80% del genoma Neandertaliano è ancora presente attualmente nell'umanità, sparso in quantità fino al 5% circa nel DNA di ogni euroasiatico.
Uomo di Denisova è la definizione provvisoria di una popolazione di Homo, le cui relazioni genetiche con le altre specie sono in fase di definizione attraverso analisi del DNA cellulare e mitocondriale.
Questi tre tipi di Homo (sapiens neandertal denisovan) più uno ulteriore non ancora individuato, come evidenziato dal team di Svante Pääbo al Max Planck Institute, si sono più volte incrociati creando un flusso genetico che ha ibridato in misure diverse ed epoche diverse tutte le popolazioni di Homo, fino ad arrivare nel nostro attuale DNA che con tecniche Archeogenetiche, che si avvalgono di supercomputers, si è potuto calcolare con sufficiente precisione la sequenza temporale e la quantità delle varie ibridazioni, fino al punto di rendersi conto che esiste un ulteriore flusso e ibridazione proveniente da un tipo di Homo che in effetti fino a ora non è mai stato ritrovato e neanche ipotizzato prima dell'avvento dell'archeogenetica di Pääbo.
In particolare si ritrovano evidenze di componenti genetiche di Homo denisova nelle popolazioni melanesiane ed asiatiche in misura minore, e forti componenti di Homo neandertal nelle popolazioni asiatiche e in misura minore europee. Si ipotizzano importanti coinvolgimenti di queste sequenze nel miglioramento del metabolismo, adattabilità e sistema immunitario, in particolare dei geni HLA di classe I, il complesso maggiore di istocompatibilità umano.
Evoluzione culturale
Si sono osservate nelle popolazioni umane in età contemporanea, in alcuni casi tendenze microevolutive superiori a quelle attese sotto l'esclusivo effetto della sola deriva genetica casuale, se presenti fattori favorenti la selezione naturale. Nell'esempio specifico un anticipo dell'età di riproduzione (età media al primo figlio scesa da 26 a 22 anni) dall'inizio del XIX secolo al 1940, in un'isola canadese a un centinaio di chilometri da Québec, in Canada. Pur con le tolleranze dovute ai fattori socioeconomici confondenti, il dato risulterebbe significativo.
L'irrilevanza, su scala temporale storica, del processo di evoluzione biologica non segna una stasi nel progresso della specie.
Pur rimanendo biologicamente la stessa specie da circa duecentomila anni, e non subendo alcuna mutazione significativa da almeno diecimila, il processo evolutivo passa all'ambito sociale, tecnologico e culturale.
Le prime e significative fasi dello stesso, in parte portate avanti da progenitori evolutivi della specie attuale, rientrano nello studio della preistoria.
La stasi apparente nell'evoluzione biologica, in realtà è un fatto dovuto alla periodizzazione considerata, nella scala temporale di riferimento.
Rarefatte esplosioni evolutive su scale a lungo termine (centinaia di migliaia/milioni di anni), d'altronde, sembrerebbero riflettere cambiamenti permanenti, come appunto la genesi di nuove specie, mentre fluttuazioni a breve termine rappresenterebbero le variazioni di nicchia locali, ottimali, selezionate da limitate variazioni ambientali all'interno di una zona, e utili a creare popolazioni all'interno della specie, adattate alle nuove condizioni.
L'apprendimento sociale diventa quindi essenziale e primario per l'adattamento umano all'ambiente, e principale motore del presente, osservabile, processo evolutivo.
Popolamento del pianeta
Esistono teorie più o meno condivise sulle origini dell'uomo contemporaneo. Esse riguardano il rapporto tra gli uomini moderni e gli altri ominidi.
La prima ondata.
- L'origine africana dell'Homo sapiens è il modello paleoantropologico dominante tra le teorie che tendono a descrivere l'origine e le prime migrazioni umane. È conosciuta anche come: ipotesi dell'origine unica, fuori dall'Africa, ipotesi africana, teoria della migrazione dall'Africa (dall'inglese Out of Africa o Out of Africa 2), origine africana recente, ipotesi di un'origine unica e recente (RSOH dall'inglese recent single-origin hypothesis), teoria del rimpiazzo (dall'inglese Replacement Hypothesis).
L'ipotesi dell'origine unica, propone che gli uomini moderni si siano evoluti in Africa e che siano poi migrati all'esterno sostituendo quegli ominidi che erano in altre parti del mondo.
Su di essa sussistono vastissime evidenze paleoantropologiche, date da diverse migliaia di ritrovamenti fossili, archeologiche, linguistiche, climatologiche (modificazioni climatiche e conseguenti selezioni della popolazione), genetiche (mtDNA e nucleare, in particolare Aplogruppi del cromosoma Y).
-- I dati molecolari condotti mediante marcatori non ricombinanti, come il DNA mitocondriale, sostengono questa ipotesi. L'analisi filo-geografica ha infatti mostrato che il popolamento da parte dell'uomo moderno dei continenti è proceduto a ondate successive a partire dal continente africano.
-- I dati paleo-antropologici testimoniano che in principio sarebbe emersa una popolazione del sud dell'Africa, isolatasi dal resto della popolazione, in un'epoca compresa tra i 100 e i 200.000 anni fa, costituita da individui piuttosto gracili rispetto ai tipi prevalenti di Homo heidelbergensis ed Homo neanderthalensis. Gli individui originari della umanità odierna furono costituiti da una popolazione Khoisan ancestrale da cui derivò l'attuale tipo etnico Khoisan. Erano individui piccoli e snelli, con una scatola cranica grande, un apparato masticatorio meno massiccio di altre specie, e presumibilmente la pelle scoperta da peli e un linguaggio evoluto. Probabilmente in origine adattati per vivere presso ambienti acquatici e di foresta in zone tropicali.
Da questo nucleo originario sarebbero derivate tre ramificazioni che dettero origine una al tipo khoisan l'altra al tipo dei pigmei africano e il terzo tipo da cui derivarono tutti gli altri tipi etnici umani. Dal terzo tipo derivò inizialmente la popolazione australoide che si diffuse partendo dall'Africa orientale circa 60.000 anni fa, e forse più, colonizzando tutta la zona tropicale fino al continente australiano, una popolazione dalla pelle scura che non utilizzava abbigliamento per difendersi dal freddo e che si diffuse seguendo le coste dei mari in cerca di molluschi. Recenti aggiornamenti basati su repertazioni in territorio arabico anticipano di alcune migliaia di anni questa fase migratoria, ipotizzando un corridoio passante per la parte meridionale del Mar Rosso, e sollevando dei dubbi sul posizionamento precedente o seguente il grande evento di riduzione della popolazione umana concomitante (Teoria della catastrofe di Toba. La teoria della catastrofe di Toba sostiene che tra 75.000 e 70.000 anni fa l'esplosione di un supervulcano al di sotto del Lago Toba, probabilmente il più grande evento eruttivo negli ultimi 25 milioni di anni, rese ancora più rigido il clima del pianeta che già stava attraversando una glaciazione).
Nel 2015 la rivista Nature ha pubblicato la notizia che alcuni denti fossili ritrovati in Cina risalirebbero a circa 120.000 anni; tale datazione sposterebbe indietro la data delle prime ondate migratorie dall'Africa, consentendo inoltre di ipotizzare che la prima colonizzazione sia avvenuta verso l'Asia.
La seconda ondata è quella del tipo dell'uomo di Cromagnon, partita dal medio-oriente circa 40.000 anni fa, che colonizzò l'Europa. Questa popolazione era di alta statura, aveva la carnagione più chiara, era dedita alla caccia di grande selvaggina, utilizzava una sofisticata tecnologia della pietra e usava pellicce per coprirsi. I resti più antichi di Homo Sapiens in Europa sono datati a 44.000 anni fa e si riferiscono a dentature rinvenute nella Grotta del Cavallo nella Baia di Uluzzo nel Comune di Nardò. I reperti, ritrovati durante scavi condotti dal prof. Palma Di Cesnola dell'Università di Siena nel 1964, finora ritenuti appartenenti all'uomo di Neandertal, sono stati oggetti di nuovo studio nel 2011: i fossili degli strati coevi alle dentature (conchiglie) esaminati al radiocarbonio nei laboratori dell'Università di Oxford per conto del Dipartimento di Antropologia dell'Università di Vienna e lo studio morfologico dello smalto delle dentature, ne hanno confermato l'appartenenza all'Homo Sapiens, spostando di almeno 4.000 anni la datazione sulla presenza dell'uomo moderno in Europa e confermando, altresì, la coabitazione almeno nell'ambito del sito del ritrovamento dell'uomo di Neandertal con l'uomo moderno.
La terza ondata (circa 25 000 anni fa) partì sempre dal medio oriente e si spinse attraverso l'Asia centrale fino in America settentrionale; di essa resta la popolazione Ainu del Giappone come esempio puro e da questa popolazione più diffusa derivarono tutti i tipi etnici successivi. La comparsa degli uomini grandi cacciatori determinò l'estinzione di molte specie animali alla fine del pleistocene, tra cui anche la scomparsa di tutte le altre specie del genere Homo.
Circa a 12.000 anni fa risale la comparsa del tipo paleomongolico che si diffuse dall'Asia centrale fino a colonizzare tutto il continente America. Circa a 7.000 anni fa risalgono le diffusioni dei tipi etnici mediterranei che dal medio oriente si diffusero in Europa e India, ed il tipo neomongolico che si diffuse in Asia orientale; questi ultimi tipi umani sono dediti alle attività economiche dell'agricoltura e allevamento. Infine vi sono le grandi diffusioni umane dei periodi storici che specie negli ultimi secoli ha portato alla grande diffusione del tipo europoide, come tipo ibrido tra i tipi Cro Magnon e mediterraneo e neo-mongolide, che attraverso la colonizzazione ha dato origine ad altre popolazioni ibride in tutti i continenti, un processo che continua oggigiorno e tende a una completa ibridazione di tutti i tipi umani.
L'ipotesi multiregionale, o della continuità regionale invece, propone che gli uomini moderni si siano evoluti, almeno in parte, da popolazioni di ominidi indipendenti.
L'ipotesi dell'origine euroasiatica o "Out of Eurasia" è una terza ipotesi alternativa, variante della multiregionale. Rivedendo in qualche modo l'ipotesi multiregionale sulla base di ritrovamenti archeologici euroasiatici, dallo studio del DNA (aplogruppo M-N-R dell'mtDNA e gli aplogruppi D-E-C-F del cromosoma Y) ipotizza un'origine euroasiatica dell'Homo sapiens.
Evoluzione tecnologica e sociale
L'agricoltura è stata una tappa delle diverse strade percorse nella civilizzazione umana.
L'uomo moderno è caratterizzato dal suo modello culturale connesso strettamente alla società a cui appartiene. Questa prospettiva, non si pone come unicità del regno animale. Viene considerato fatto acquisito dalle ricerche in ambito neurobiologico e neurocognitivista che l'espressione culturale non abbia assolutamente un valore extra-naturale. Inoltre altri animali (delfini, primati, elefanti, ecc) presentano un passaggio di conoscenza in senso orizzontale, presentano quindi cultura. È evidente però che nell'essere umano questa attitudine è ipertrofica, se paragonata ad altri animali.
Dalla sua comparsa sulla terra fino ad oggi, culture e società umane hanno continuamente subito mutamenti, in un lento e progressivo sviluppo che ha portato a un'evoluzione culturale dell'essere umano, chiamata sinteticamente progresso.
Nel corso della storia le varie società si sono sviluppate, portando avanti una complessità sempre crescente, con l'aumentare delle conoscenze, delle varie culture e delle popolazioni. Crisi alimentari, scoperte e rivoluzioni scientifiche e sociali hanno spesso segnato i passaggi di questa evoluzione.
Il progresso della civiltà umana ha portato al prolungamento della speranza di vita e al miglioramento delle condizioni igienico sanitarie, al riconoscimento di diritti dell'uomo, alla maggior conoscenza delle risorse naturali e al miglioramento della qualità di vita generale, anche se a tutt'oggi in molte parti del mondo queste evoluzioni sono avvenute solo in parte o non sono ancora potute avvenire.
Caratteristica di questa parte evolutiva è la sempre maggiore velocità nel trasferimento orizzontale di elementi culturali, reso sempre più efficiente dai mezzi tecnologici che, a partire dalla scrittura, hanno reso possibile il fenomeno. La trasmissione veloce, a distanza, delle informazioni a partire dal XIX secolo, ha velocizzato il fenomeno, cresciuto esponenzialmente nel XX e XXI con le tecnologie digitali.
Un aspetto interessante, e che gli studiosi cerano di comprendere, è come mai il cro-magnum avesse una capacità cranica di circa 1700 cm3, il neandertaliano di circa 1800 cm3, entrambi superiore ai circa 1200 dell'homo sapiens, eppure, questo sia riuscito a prevalere sugli altri due.
LE ORIGINI DELL'UOMO
LE PRIME IMPRONTE
NASCITA DELLA COPPIA
IL PRIMO OMINIDE FOSSILE
L'AUSTRALOPITECO DIVENTA ERBIVERO
INIZIA L'ETA' DELLA PIETRA
HOMO ERGASTER
HOMO ERECTUS
IL CERVELLO E LO STRUMENTO
L'UOMO DI CEPRANO
L'ENIGMA DI ISERNIA
LA CONQUISTA DEL FUOCO
LA SCOPERTA DI ATAPUERACA
L'UOMO DI SACCOPASTORE
I NEANDERTAL
HOMO SAPIENS
Eugenio Caruso -15 - 02- 2022

