Da Lucy al genere Homo, fossili e DNA stanno ricomponendo il cespuglio dellevoluzione umana
Il punto sulla nostra
storia evolutiva
Un secolo fa,
al tempo in cui infuriava il dibattito
scientifico sull«anello mancante» dellevoluzione
umana, una studentessa di nome
Josephine Salmons attirò lattenzione
di Raymond Dart, professore allUniversità
del Witwatersrand in Sudafrica, su un
cranio fossile che aveva visto a casa di un
amico.
Interessato a fondare una collezione
di fossili nellateneo, Dart si recò sul sito
del ritrovamento, dove nel giro di poche
settimane scoprì un altro cranio, che illustrò
su «Nature» allinizio del 1925. Il bambino
di Taung, che Dart denominò Australopithecus
africanus, aveva tutta laria di
essere un antenato umano.
Nelle parole di
Dart, un «anello di congiunzione estinto
tra luomo e il suo antenato scimmiesco».
Se il bambino di Taung, datato a 2,8 milioni
di anni fa, avrebbe sconvolto gli studi
sulle origini delluomo, supportando lipotesi
di Darwin che lAfrica fosse la culla
dellumanità, 35 anni più tardi nella Gola di
Olduvai, in Tanzania, Louis e Mary Leakey
scoprirono un fossile che sembrava individuare
la più antica specie del nostro genere.
Battezzata Homo habilis e risalente a 1,8
milioni di anni fa, oggi la sua classificazione
è ancora controversa, per via di una serie di
caratteristiche ancora molto primitive.
Anni dopo, Maurice Taieb riunì un nutrito
gruppo di paleoantropologi di fama
mondiale, tra cui Donald Johanson e Yves
Coppens, per aprire uno scavo nel promettente
sito di Hadar, nella Depressione
dellAfar, in Etiopia.
Alla seconda stagione
di scavi, il 24 novembre 1974 Johanson
si imbatté nello scheletro incredibilmente
ben conservato, risalente a 3,2 milioni
di anni fa, di un esemplare di una nuova
specie, che avrebbero poi chiamato Australopithecus
afarensis. Il reperto fu ufficialmente
catalogato con la sigla A.L. 288-1
ma, si sa, i paleoantropologi amano dare
nomignoli alle loro creature, e nel campo
risuonavano di continuo le note di una celebre
canzone dei Beatles. E Lucy fu.
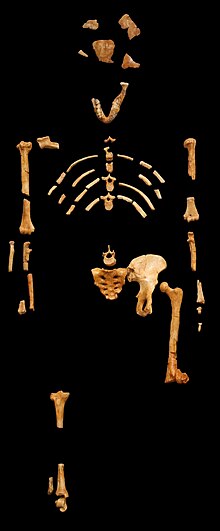
Ricostruzione dello scheletro di Lucy, esposta presso il Museo nazionale di storia naturale di Francia, a Parigi.
Sarà stato il nomignolo, o labbondanza
di ossa che permettevano di ricostruirne
quasi completamente lo scheletro, sarà
stato lentusiasmo degli addetti ai lavori per
quello che sembrava essere il nostro antenato
più antico, fatto sta che Lucy divenne
subito una star mediatica.
A maggior ragione
perché lanalisi delle sue caratteristiche
anatomiche portava a dedurre che camminasse
con postura eretta.
Una conclusione
che avrebbe trovato conferma nel 1978,
quando ancora i coniugi Leakey trovarono
a Laetoli, in Tanzania, le famose impronte
fossili risalenti a 3,7 milioni di anni fa e attribuite
alla stessa specie, A. afarensis.
Tutti questi compleanni sono loccasione
per fare il punto su quello che abbiamo
scoperto di nuovo sulla nostra storia,
su come si stia ricomponendo il cespuglio
genealogico dellevoluzione umana grazie
a nuovi fossili e nuovi strumenti di analisi,
compreso lo studio dei DNA antichi, fin
dove può esserci di aiuto.
Se potessimo tornare indietro nel tempo a circa 6
milioni di anni fa, quando comparvero in Africa
le prime forme di ominini, il gruppo di specie di
cui facciamo parte noi e i nostri antenati, sarebbe
impossibile immaginare il percorso evolutivo
che ha portato, milioni di anni dopo, alla comparsa
della nostra specie, Homo sapiens. Levoluzione
ha agito attraverso complesse dinamiche di adattamenti
locali, variazioni casuali, estinzioni e ripopolamenti, dettati da
condizioni contingenti difficilmente prevedibili. A posteriori, tuttavia,
è possibile individuare in questo percorso alcuni momenti
chiave che segnano fasi di transizione e di cambiamento concentrate
nel tempo geologico.
Una di queste serie di eventi chiave è quella che si è verificata
nelle fasi finali del Pliocene e che ha portato alla comparsa delle più
antiche specie attribuite al genere Homo. Nellarco temporale tra 4
e 3 milioni di anni fa i resti fossili di ominini sono relativamente numerosi,
con varie specie fossili descritte e assegnate in larga misura
al genere Australopithecus. A partire da circa 3 milioni di anni fa,
sotto la spinta di un progressivo inaridimento del clima e di un aumento
della variabilità nella successione di periodi umidi e periodi
aridi, queste specie vanno incontro a pressioni selettive che hanno
come risultato lorigine di due gruppi, profondamente diversi
dal punto di vista morfologico dalle forme precedenti e diversi anche
tra loro; un evento che viene descritto con il termine «bivio
adattativo».
Le specie di questi due gruppi sono di solito attribuite
a due generi diversi, il genere Paranthropus e il genere Homo.
Le specie di Paranthropus sono caratterizzate da una specializzazione
dellapparato masticatorio, con aumento delle dimensioni
dei denti premolari e molari, probabilmente legata alla necessità
di elaborare cibi molto fibrosi. Queste specie sono documentate
in Africa orientale e in Sudafrica fino a circa un milione di anni fa,
quando si sono estinte. Laltro gruppo è quello del genere Homo.
Le prime scoperte di quella finora considerata la più antica
specie del genere Homo (H. habilis) risalgono agli anni sessanta
quando, in livelli pleistocenici nella Gola di Olduvai, in Tanzania
i paleoantropologi Louis e Mary Leakey scoprirono un fossile oggi
indicato con la sigla OH 7 (cioè Olduvai Hominid 7), designato
poi come lolotipo (termine che indica il reperto sul quale viene
descritta una specie) della specie Homo habilis, rappresentato da
vari elementi appartenenti a un individuo subadulto, fra cui due
ossa del cranio, una mandibola e 13 ossa delle mani.
Le ossa parietali del cranio, pur se incomplete, consentirono di
stimare la capacità cranica a circa 650 centimetri cubi. Questa stima,
superiore a quella dei reperti di Australopithecus noti fino a
quel momento, insieme alle dimensioni relativamente ridotte dei
denti mandibolari, convinse i ricercatori che si trovavano davanti
a una specie di ominino più avanzata rispetto ad Australopithecus.
Per questo nel 1964 i paleoantropologi Louis Leakey, Phillip Tobias
e John Napier decisero di definire una nuova specie e attribuirla
al genere Homo, chiamandola Homo habilis, per le dimensioni
cerebrali ricostruite ma anche per la presenza, negli stessi livelli
dei fossili, di semplici strumenti in pietra, da allora indicati come
utensili di tipo olduvaiano. Analisi radiometriche successive portarono
poi a definire letà dei fossili a circa 1,8 milioni di anni fa.
Negli anni seguenti altri fossili simili ritrovati a Olduvai confermarono
la presenza di questa specie. Si venne così a creare un paradigma
che legava la presenza di strumenti in pietra (che faceva
supporre una notevole capacità manipolativa) a elevate dimensioni
cerebrali, entrambi elementi che sembravano ben definire le
più antiche specie del genere Homo.
Come spesso succede in paleoantropologia, la descrizione della
specie H. habilis venne accolta con scetticismo. Prima di
H. habilis la più antica specie del genere Homo che era stata proposta
era Homo erectus, descritta alla fine dellOttocento sulla base
di resti ritrovati in Indonesia, e si riteneva che non potesse essere
esistita alcuna forma di Homo più arcaica di H. erectus. Molti
autori sostenevano che i nuovi fossili da Olduvai non fossero così
dissimili da quelli della specie Australopithecus africanus trovati
in Sudafrica nei decenni precedenti, per cui le forme di Olduvai
dovevano rappresentare una specie di Australopithecus tipica
dellAfrica orientale. Poi, allinizio degli anni settanta, nuove scoperte
in un sito nel nord del Kenya denominato Koobi Fora, sulla
sponda est del Lago Turkana, sembrarono dar ragione agli autori
che avevano designato la specie H. habilis.

Lo scheletro di H. ergaster KNM-WT 15000, il «ragazzo del Turkana
». Il cranio è completo a parte i tori sopraorbitari; lo scheletro assiale
presenta buona parte delle ossa, almeno da un lato. Mancano
la maggior parte delle vertebre cervicali e alcune vertebre toraciche,
parti del sacro e dellosso iliaco e alcune coste. Gli arti superiori
sono privi della scapola e dellomero sinistri, di entrambi i radii
e della maggior parte delle ossa della mano. Gli arti inferiori sono
privi dellestremità prossimale del femore destro, della maggior
parte delle epifisi delle ossa della gamba e della maggior parte delle
ossa del piede. Il volume endocranico è di 909 centimetri cubi.
Le ricerche a Koobi Fora erano guidate da Richard Leakey (figlio
di Louis e Mary) e dalla moglie Meave. Nellarco di pochi anni
venne alla luce unenorme quantità di fossili di ominini, poi attribuiti
in parte a Paranthropus boisei e in parte a H. habilis, sulla
base di affinità morfologiche con i fossili di Olduvai. Fra i reperti
più significativi vi sono due crani, relativamente ben conservati,
ai quali ci si riferisce con il loro numero di catalogo del Kenya
National Museum, e cioè KNM-ER 1470 (datato a circa 2,1 milioni
di anni fa) e KNM-ER 1813 (1,9) . Le ridotte dimensioni dentarie e lelevata capacità cranica
furono fra gli elementi che indussero Richard Leakey ad attribuire
questi fossili alla specie H. habilis. Le notevoli differenze fra i due
nelle dimensioni complessive vennero interpretate come dovute
a dimorfismo sessuale: KNM-ER 1470 fu considerato un individuo
maschile e KNM-ER 1813 uno femminile. I due fossili sembravano
quindi confermare la validità della specie H. habilis, che venne
progressivamente riconosciuta dalla maggior parte degli autori.
In questa formidabile collezione di resti fossili vi sono altri due
reperti molto significativi. Il primo è un altro cranio ben conservato,
noto come KNM-ER 3733. Il secondo reperto è uno spettacolare scheletro quasi
completo di un individuo non ancora adulto indicato come KNMWT
15000, noto anche come il ragazzo del Turkana, proveniente
dalla località di Nariokotome, sulla
sponda occidentale del lago Turkana
e datato a circa 1,5 milioni di anni fa.
Lo studio dei resti craniali, mandibolari
e dentari di Koobi Fora fu condotto
da Bernard Wood, ora alla George Washington University,
che, con un accurato lavoro di descrizione e analisi metrica comparativa,
concluse che nel campione di Koobi Fora potevano essere
individuate tre specie diverse appartenenti al genere Homo.
Una era riconducibile a H. habilis e rappresentata dal cranio KNMER
1813, mentre le altre due erano rappresentate rispettivamente
dai crani KNM-ER 1470 e KNM-ER 3733, che mostravano una
serie di caratteri diversi rispetto a quelli di H. habilis. Alla luce di ciò, Wood propose nel
1992 il nome di H. rudolfensis per la specie rappresentata dal cranio
KNM-ER 1470 e quello di H. ergaster per i reperti come KNMER
3733 e il ragazzo del Turkana. Forme analoghe a H. ergaster (indicate
come Homo aff. erectus, modalità con cui si indicano fossili
che presentano affinità con H. erectus) si ritrovano anche nei siti
sudafricani di Swartkrans, Drimolen e Kromdraai, con unantichità
simile o maggiore di quella dei siti dellAfrica orientale, anche se
la documentazione fossile è più frammentaria.
Se la specie H. ergaster fu riconosciuta valida da molti autori
(mentre altri la considerano la variante africana della specie
H. erectus, distribuita anche in Asia), la definizione di H. rudolfensis
come specie distinta da H. habilis lasciò perplessi i ricercatori,
che interpretavano le differenze tra i due gruppi di fossili come
dovute alla variabilità intraspecifica, quindi anche in termini
di dimorfismo sessuale. Poi, nel 2012, la descrizione e linterpretazione
di nuovi reperti craniali rinvenuti nella località di Ileret
anchessa sulle sponde del lago Turkana, ha indicato le affinità di
questi col cranio KNM-ER 1470 e le differenze rispetto al cranio
KNM-ER 1813, portando quindi alla loro attribuzione a H. rudolfensis
e confermando lipotesi di Wood del 1992.
Oggi si viene quindi a delineare lesistenza di una radiazione
adattativa alle origini del genere Homo, con tre specie diverse e
in parte coeve: H. habilis, i cui fossili coprono un intervallo temporale
tra 2,1 e 1,65 milioni di anni fa; H. rudolfensis (2,1-1,78) e H.
ergaster (2,0-1,0). Levidenza fossile indica anche che alcune popolazioni
di queste specie sono coesistite negli stessi areali, come
larea sulle sponde nel nord-est del Lago Turkana. Si può quindi
pensare che avessero strategie adattative differenti tra loro per
poter occupare gli stessi ambienti.
La distribuzione temporale di tutte e tre queste specie a partire
da 2 milioni di anni fa suggerisce anche che il modello tradizionale
di una semplice derivazione di H. ergaster/erectus da H. habilis
è probabilmente troppo semplicistico. Inoltre, la presenza di
tre specie diverse fra loro morfologicamente già a 2 milioni di anni
fa prefigura unorigine ben più antica del genere Homo.
A questo
riguardo i resti fossili attribuiti a Homo nellarco temporale fra
i 3 e i 2 milioni di anni fa si contano sulle dita di una mano. A parte
i ritrovamenti di singoli denti isolati in alcune località dellAfrica
orientale, le evidenze più complete sono relative a tre soli fossili.
Si tratta di reperti isolati, distribuiti su un ampio arco temporale
e su una vasta area geografica: una mandibola completa con
denti, scoperta nel sito di Uraha, in Malawi, datata fra i 2,3 e i 2,5
milioni di anni fa e attribuita a Homo rudolfensis; una mascella con
alcuni denti scoperta nel sito di Hadar in Etiopia, datata a 2,33 milioni
di anni fa e attribuita a Homo aff. habilis; e una emi-mandibola
sinistra con alcuni denti premolari e molari, rinvenuta nella località
di Ledi Geraru, in Etiopia, e datata a circa 2,8 milioni di anni
fa, senza unattribuzione specifica. La combinazione di tratti morfologici
primitivi già presenti in Australopithecus con tratti più derivati
tipici delle specie di Homo più recenti permette di considerare
questi fossili come le più antiche evidenze del genere Homo.
La mandibola di Ledi Geraru è di particolare interesse in quanto
rappresenta a oggi il più antico fossile di una specie di Homo e
testimonia una lunga storia evolutiva, che precede di molto le evidenze
che troviamo a partire da circa 2 milioni di anni fa; unidea
che deriva anche da un recente lavoro di rianalisi dellolotipo di
H. habilis OH 7. Resta da stabilire quali siano le precise relazioni
tra questi tre fossili e le specie successive. Pur nella notevole diversità
morfologica, le tre specie sono tutte caratterizzate da una
capacità cranica media più grande di quella di Australopithecus,
una maggiore altezza della volta cranica, un prognatismo facciale
ridotto, denti premolari e molari più stretti e lunghi.

Ledi-Geraru è un sito archeologico che si trova lungo i fiumi Ledi e Geraru, due affluenti di sinistra dell'Auasc, nella regione degli Afar in Etiopia.
Le prime ricerche furono condotte tra il 1972 e il 1974. Il sito è noto soprattutto perché qui, nel 2013, fu trovata una mandibola di ominide, un fossile repertato come LD 350-1, datata 2,8 milioni di anni, che potrebbe rappresentare uno dei primi esemplari del genere Homo.
Qui sono stati ritrovati anche diversti strumenti in pietra, datati circa 2,6 milioni di anni, come anche ossa di animali, appartenute ad animali come gazzelle e giraffe, tipiche delle savane.
Linterpretazione della diversità morfologica nei fossili attribuiti
a H. habilis, H. rudolfensis e H. ergaster è stata oggetto di molti
lavori per cercare di stabilire le relazioni tra le varie specie. Da
metà anni novanta vari specialisti condussero analisi filogenetiche
usando le nuove specie descritte come gruppi distinti (tecnicamente
detti unità tassonomiche), e confrontandole sia con specie
di Australopithecus sia con varie specie di Homo. Pubblicazioni
diverse portavano a risultati differenti circa le affinità di H. habilis,
H. rudolfensis e H. ergaster con Australopithecus, H. erectus e
H. sapiens. Il gruppo delle prime tre specie non appariva come un
gruppo omogeneo, al di là di vaghe affinità morfologiche quali una
capacità cranica assoluta superiore a quella delle specie di Australopithecus.
La questione quindi si pose in questi termini: a parte le
differenze fra loro a livello specifico, quali sono gli elementi che
distinguono queste specie nel loro complesso da quelle del genere
Australopithecus? O anche, quali sono i caratteri che permettono
la loro inclusione nel genere Homo? E, più in generale, quali sono i
caratteri che definiscono il genere Homo? Il tema era di importanza
notevole, dato che riguardava anche la definizione della nostra
specie genere Homo, specie H. sapiens e fu affrontato nel 1999
da Wood e Mark Collard, allora allo University College di Londra,
in un significativo lavoro di sintesi, aggiornato poi nel 2015 sulla
base di nuove ricerche e nuovi fossili scoperti dopo il 1999.
In primo luogo, Wood e Collard partirono dalla definizione
della categoria tassonomica di «genere» in biologia. Rivedendo il
concetto di genere definito da Ernst Mayr nel 1950, proposero che
un genere deve essere un gruppo di specie (o anche una sola specie)
monofiletico, cioè derivante da un unico antenato, che condivide
una singola zona adattativa. Questo termine indica linsieme
delle strategie adattative che le varie specie mettono in atto per
sopravvivere in un dato ambiente, quali procurarsi il cibo o riprodursi.
Circa alcune di queste strategie è possibile ricavare informazioni
dalla documentazione paleontologica; per esempio le dimensioni
corporee, le modalità di locomozione, le caratteristiche
dellapparato masticatorio, eccetera.
Per poter stabilire se un gruppo di specie è assegnato correttamente
a un determinato genere, Wood e Collard proposero
due criteri:
1) le specie devono appartenere allo stesso gruppo
monofiletico della «specie tipo» di quel genere, dove nel caso del
genere Homo la specie tipo è H. sapiens, dato che è quella che è
stata descritta per prima (nel nostro caso da Linneo nel 1758);
2) le
strategie adattative delle specie di un genere devono essere più simili
a quelle della specie tipo del genere nel quale sono incluse rispetto
a quelle della specie tipo di un altro genere, per esempio A.
africanus, specie tipo del genere Australopithecus.
Un esempio permette di chiarire il concetto. Uno dei caratteri
distintivi della nostra specie è landatura bipede obbligata, cioè
esclusiva (non siamo in grado di muoverci agevolmente usando
altre modalità di locomozione, quali larrampicamento o landatura
quadrupede). Quindi landatura bipede obbligata è uno degli
elementi che deve caratterizzare tutte le specie che appartengono
al genere Homo.
Il primo passo del lavoro di Wood e Collard fu stabilire se le specie
appartenenti al genere Homo formassero un gruppo monofiletico
o meno. Le specie considerate furono H. sapiens, H. neanderthalensis,
H. heidelbergensis, H. erectus, H. ergaster, H. habilis,
H. rudolfensis, oltre a quelle dei generi Australopithecus e Paranthropus.
Attraverso una sofisticata analisi filogenetica, Wood
e Collard misero in evidenza che soltanto le specie H. sapiens, H.
neanderthalensis, H. heidelbergensis, H. erectus e H. ergaster formavano
un gruppo monofiletico ben definito. Le relazioni filogenetiche
di H. habilis e H. rudolfensis risultavano più ambigue.
Per il secondo criterio, linsieme delle strategie adattative dello
stesso gruppo di cinque specie (cioè le informazioni derivanti dalla
ricostruzione di dimensioni e forma del corpo, modalità di locomozione
e tipo di dieta) risultò più simile a quello di H. sapiens
che non di A. africanus; per H. habilis e H. rudolfensis, invece, le
evidenze mostravano maggiori affinità con Australopithecus.
Le conclusioni di Wood e Collard furono quindi che, per rispettare
i criteri stabiliti per linclusione nel genere Homo, le specie
H. habilis e H. rudolfensis dovrebbero essere rimosse da questo
genere e incluse, come soluzione più semplice, nel genere
Australopithecus. Quindi, alla luce delle ipotesi di Wood e Collard,
la più antica specie del genere Homo sarebbe H. ergaster (da alcuni
indicato come H. erectus africano) la cui struttura scheletrica,
ben documentata dal reperto del ragazzo del Turkana, presenta
una «architettura» sostanzialmente di tipo moderno, che dimostra
un adattamento allandatura bipede obbligata e alla corsa di
resistenza.
Lidea che le specie H. habilis e H. rudolfensis siano più simili
a forme di Australopithecus che non di Homo ha trovato negli
ultimi anni più conferme, derivanti da una serie di ricerche indipendenti
fra loro su temi diversi relativi alle strategie adattative di
queste e delle altre specie di ominini. In breve, queste evidenze
riguardano: 1) la locomozione; 2) il tipo di dieta; 3) la struttura interna
dei denti.
Per le indicazioni relative alla modalità di locomozione il fossile
chiave è rappresentato da uno scheletro parziale scoperto a
Olduvai e descritto nel 1987, indicato come OH 62 e attribuito a H.
habilis, lunico scheletro relativamente completo di questa specie.
Il reperto ha losso del braccio (omero) relativamente più lungo
dellosso della coscia (femore). Già questa era una prima indicazione
del fatto che questo esemplare avesse proporzioni degli arti
più simili a quelle di Australopithecus che di Homo. Poi nel 2009
Chris Ruff, della Johns Hopkins University, analizzò lo spessore
della sezione più esterna dellomero e del femore (porzione indicata
come corticale), confrontandola con la condizione osservabile
in H. sapiens e negli scimpanzé. La differenza principale tra
noi e gli scimpanzé sta nel fatto che in questi ultimi lo spessore
della corticale dellomero è molto più marcato, e ciò rappresenta
un adattamento per resistere alle forze che si generano quando le
braccia vengono usate per arrampicarsi sugli alberi. In H. sapiens
lo spessore della corticale dellomero è minore, perché con landatura
bipede obbligata luso dellarto superiore è ridotto, in quanto
non lo si impiega per arrampicarsi sugli alberi. In OH 62 la struttura ossea dellomero rispetto al femore è molto simile a quella
degli scimpanzé, a indicare come questo individuo (e quindi per
estensione la specie H. habilis) avesse ancora una forte componente
di arrampicamento nel proprio repertorio di locomozione.
Si tratta in questo caso di unevidenza relativa non solo alla morfologia
esterna (le proporzioni degli arti) ma anche alla funzionalità
dellosso, che dimostra una condizione analoga ad Australopithecus
e non alle altre specie del genere Homo.
Una tecnica che consente di ricostruire la dieta dei Mammiferi
(viventi ed estinti) prevede di analizzare il contenuto di alcuni
elementi chimici presenti nelle ossa e nei denti, e in particolare
le loro forme isotopiche stabili. Gli isotopi usati in questo tipo di
analisi sono di solito il carbonio-13 e lazoto-15, che si sono rivelati
buoni indicatori del tipo di dieta, sia negli animali sia in varie specie
di ominini fossili, e anche in popolazioni di H. sapiens. Questa
analisi, e in particolare lo studio del carbonio-13 nelle ossa, è stata
condotta nel 2019 da David Patterson della University of North
Georgia e colleghi, su un campione molto ampio di specie fossili
di mammiferi africani rinvenuti in vari siti dellAfrica orientale
nellarco di tempo tra 4,4 e 1,5 milioni di anni fa. Fra le specie considerate
vi erano anche otto specie di ominini, dei generi Ardipithecus,
Australopithecus, Paranthropus e Homo. Il dato che emerge
tecnicamente indicato come «segnale isotopico» dei livelli
di carbonio-13 è molto simile nelle specie più antiche quali Ardipithecus
ramidus e poi Australopithecus afarensis, fino a H. habilis
e H. rudolfensis inclusi. Questo indica una dieta in larga misura
frugivora per questo gruppo di specie.
La cesura nel segnale
isotopico avviene nelle specie più recenti rispetto a H. habilis, a
iniziare da H. ergaster, e ciò è stato interpretato come legato a un
marcato cambiamento di dieta; questo sarebbe da ricondurre a
una maggiore componente carnea nella dieta di H. ergaster legata
a un più elevato consumo di prede animali, in larga misura la
vasta gamma di specie di ungulati presenti nelle savane africane,
attivamente cacciate da questi ominini. Il cambiamento radicale
nel tipo di dieta con tutte le conseguenze che ciò ha comportato,
e cioè una dieta di più elevata qualità che avrebbe permesso
un ulteriore incremento delle dimensioni cerebrali sarebbe avvenuto
quindi a partire da H. ergaster, e non prima.
Il terzo tipo di evidenza che ha permesso di riconsiderare la
classificazione di H. habilis e H. rudolfensis deriva dallanalisi della
struttura interna dei denti molari e premolari, e in particolare
di quella zona del dente che prende il nome di giunzione smalto-
dentina. Questarea si trova allinterno di ognuno dei nostri
denti nella zona di contatto, come dice il nome, tra lo smalto, che
rappresenta lo strato più esterno che compone buona parte della
corona dentaria, e la dentina, che costituisce la struttura portante
del dente al suo interno, incluse le radici. Lo strato di cellule
che compone la giunzione smalto-dentina si forma nelle primissime
fasi dello sviluppo dentario e riflette linformazione genetica
che contribuisce alla definizione della forma finale del dente. La
sua morfologia ha una forte componente genetica e, rispetto ad
altre porzioni dello scheletro, è influenzata solo in misura minima
da fattori ambientali. Questa morfologia rimane intatta anche nel
dente adulto, e può essere analizzata e quantificata grazie a tecniche
di microtomografia computerizzata ad alta risoluzione.
Negli ultimi ventanni sono stati condotti numerosi studi sulla
morfologia della giunzione smalto-dentina in varie specie di primati
viventi e fossili, ed è emerso che questa consente di discriminare
bene fra loro i denti di specie diverse di primati anche molto
vicini filogeneticamente, quali per esempio, fra gli scimpanzé, le
due specie Pan troglodytes e Pan paniscus. Fra le specie di ominini
fossili sono state analizzate specie del genere Australopithecus,
Paranthropus e Homo, da H. erectus fino a H. neanderthalensis e
H. sapiens. Solo molto di recente, nel 2024, lanalisi della giunzione
smalto-dentina dei denti premolari e molari è stata applicata
ai reperti di H. habilis e H. rudolfensis dai siti dellAfrica orientale,
fra i quali Olduvai e Koobi Fora. I risultati sono stati abbastanza
sorprendenti, ma non inaspettati, in quanto la massima parte
di questi denti ricadono, per la loro morfologia, allinterno della
variabilità del genere Australopithecus, e ben distinti rispetto alle
altre specie del genere Homo. Il gruppo che include H. ergaster
e H. erectus, invece, ha una morfologia della giunzione smaltodentina
ben distinguibile da quella degli australopitechi. Anche
per quanto riguarda la morfologia dentaria, quindi, la transizione
verso forme diverse da Australopithecus sembra essere avvenuta
non con H. habilis e H. rudolfensis, bensì con H. ergaster.
Dal complesso delle tre linee di evidenze
discusse, del tutto indipendenti
tra loro (modalità di locomozione,
dieta e ultrastruttura dentaria), pare
emergere in maniera chiara lipotesi
che fossili finora attribuiti a H. habilis
e H. rudolfensis dovrebbero trovare una diversa collocazione
sistematica. Una possibilità potrebbe essere, come suggerivano
Wood e Collard, di considerarle come specie del genere Australopithecus;
unaltra quella di inserirli in un nuovo genere, tuttora
da definire.
Esattamente sessantanni fa, nel 1964, veniva descritta la specie
H. habilis, allora considerata la più antica specie di Homo,
da cui si riteneva si fosse originato H. erectus. Oggi il quadro che
abbiamo dellorigine del genere Homo è profondamente cambiato,
grazie a nuovi fossili descritti in questi anni e a nuove datazioni
dei contesti di provenienza. Inoltre, le recenti interpretazioni
sono anche il risultato dellimpiego di nuovi e sofisticati metodi
di analisi dei dati morfologici e di altri tipi di dati che consentono
di ricostruire le strategie adattative di queste specie. Le nuove
evidenze hanno mostrato unantichità del genere Homo di gran
lunga superiore a quanto pensavamo fino a pochi anni fa e lesistenza
di tre specie distinte (H. habilis, H. rudolfensis, H. ergaster/
erectus) fra loro coeve, a indicare una radiazione adattativa (il termine radiazione adattativa o radiazione evolutiva descrive un fenomeno di rapida (in termini geologici) diversificazione di nuove specie a partire da un progenitore comune) che risale
probabilmente alle fasi finali del Pliocene, piuttosto che una
semplice relazione di antenato/discendente tra H. habilis e H. ergaster/
erectus.
Altre questioni rimangono aperte, quali le relazioni filogenetiche
fra le tre specie e la posizione sistematica di H. habilis e H. rudolfensis
come specie del genere Homo o meno. Tutto questo ci
consente oggi di proporre nuove ipotesi e nuovi modelli evolutivi
che potranno essere ulteriormente testati in futuro, anche grazie
a nuovi fossili, ancora in attesa di essere scoperti.
di Jacopo Moggi Cecchi e Alessandro Riga
Eugenio Caruso - 23 gennaio 2025

Tratto da le scienze








